无尾感器纲
尾感器纲
你的位置:
首页
>
植物界
> 植物组织
细胞分化导致植物体中形成多种类型的细胞,这也就是细胞分化导致了组织的形成。人们一般把在个体发育中,具有相同来源的(即由同一个或同一群分生细胞生长、分化而来的)同一类型,或不同类型的细胞群组成的结构和功能单位,称为组织(tissue)。由一种类型细胞构成的组织,称简单组织(simple tissue)。由多种类型细胞构成的组织,称复合组织(compoundtissue)。植物每一类器官都包含有一定种类的组织,其中每一种组织具有一定的分布规律和行使一种主要的生理功能,但是这些组织的功能又是必须相互依赖和相互配合的,例如叶是植物进行光合作用的器官,其中主要分化为大量的同化组织进行光合作用,但在它的周围覆盖着保护组织,以防止同化组织丢失水分和机械损伤,此外,输导组织贯穿于同化组织中,保证水分的供应和把同化产物运输出去,这样,三种组织相互配合,保证了叶的光合作用正常进行。由此可见,组成器官的不同组织,表现为整体条件下的分工协作,共同保证器官功能的完成。植物组织分成分生组织(meristematic tissue)和成熟组织(mature tissue)两大类。
组织
tissue
线虫动物门的主要特征
出现原体腔(初生体腔)
消化系统有完全的消化道,有口和肛门
角质膜起保护作用,有蜕皮现象
排泄系统为原肾管,无纤毛和焰细胞,分腺型和管型
圆筒状神经系统,感官不发达
管状生殖系统,雌雄同体、异形
分生组织 meristematic tissue
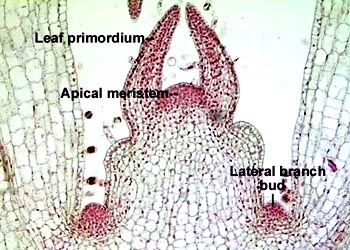
种子植物中具分裂能力的细胞限制在植物体的某些部位,这些部位的细胞在植物体的一生中持续地保持强烈的分裂能力,一方面不断增加新细胞到植物体中,另一方面自己继续“永存”下去,这种具持续分裂能力的细胞群称为分生组织。根据在植物体上的位置,可以把分生组织区分为顶端分生组织(apical meristem)、侧生分生组织(lateral meristem)和居间分生组织(intercalary meristem)。顶端分生组织位于茎与根主轴的和侧枝的顶端。它们的分裂活动可以使根和茎不断伸长,并在茎上形成侧枝和叶,使植物体扩大营养面积。茎的顶端分生组织最后还将产生生殖器官。顶端分生组织细胞的特征是:细胞小而等径,具有薄壁,细胞核位于中央并占有较大的体积,液泡小而分散,原生质浓厚,细胞内通常缺少后含物。侧生分生组织位于根和茎的侧方的周围部分,靠近器官的边缘。它包括形成层(cambium)和木栓形成层(cork cambium或phellogen)。形成层的活动能使根和茎不断增粗,以适应植物营养面积的扩大。木栓形成层的活动是使长粗的根、茎表面或受伤的器官表面形成新的保护组织。侧生分生组织并不普遍存在于所有种子植物中,它们主要存在于裸子植物和木本双子叶植物中。草本双子叶植物中的侧生分生组织只有微弱的活动或根本不存在,在单子叶植物中侧生分生组织一般不存在,因此,草本双子叶植物和单子叶植物的根和茎没有明显的增粗生长。侧生分生组织的细胞与顶端分生组织的细胞有明显的区别,例如形成层细胞大部分呈长梭形,原生质体高度液泡化,细胞质不浓厚。而且它们的分裂活动往往随季节的变化具有明显的周期性。居间分生组织是夹在多少已经分化了的组织区域之间的分生组织,它是顶端分生组织在某些器官中局部区域的保留。典型的居间分生组织存在于许多单子叶植物的茎和叶中,例如水稻、小麦等禾谷类作物,在茎的节间基部保留居间分生组织,所以当顶端分化成幼穗后,仍能借助于居间分生组织的活动,进行拔节和抽穗,使茎急剧长高。葱、蒜、韭菜的叶子剪去上部还能继续伸长,这也是因为叶基部的居间分生组织活动的结果。落花生由于雌蕊柄基部居间分生组织的活动,而能把开花后的子房推入土中。居间分生组织与顶端分生组织和侧生分生组织相比,细胞持续活动的时间较短,分裂一段时间后,所有的细胞都完全转变成成熟组织。
顶端分生组织
(apical meristem)位于茎与根主轴的和侧枝的顶端。它们的分裂活动可以使根和茎不断伸长,并在茎上形成侧枝和叶,使植物体扩大营养面积。茎的顶端分生组织最后还将产生生殖器官。顶端分生组织细胞的特征是:细胞小而等径,具有薄壁,细胞核位于中央并占有较大的体积,液泡小而分散,原生质浓厚,细胞内通常缺少后含物。
侧生分生组织
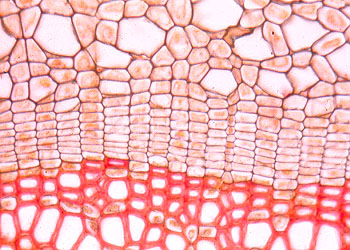
(lateral meristem)位于根和茎的侧方的周围部分,靠近器官的边缘。它包括形成层和木栓形成层。形成层的活动能使根和茎不断增粗,以适应植物营养面积的扩大。木栓形成层的活动是使长粗的根、茎表面或受伤的器官表面形成新的保护组织。侧生分生组织并不普遍存在于所有种子植物中,它们主要存在于裸子植物和木本双子叶植物中。
居间分生组织
(intercalary meristem)是夹在多少已经分化了的组织区域之间的分生组织,它是顶端分生组织在某些器官中局部区域的保留。典型的居间分生组织存在于许多单子叶植物的茎和叶中,例如水稻、小麦等禾谷类作物,在茎的节间基部保留居间分生组织,所以当顶端分化成幼穗后,仍能借助于居间分生组织的活动,进行拔节和抽穗,使茎急剧长高。
分生组织也可根据组织来源的性质划分为原分生组织(promeristem)、初生分生组织(primary meristem)和次生分生组织(secondary meristem)。原分生组织是直接由胚细胞保留下来的,一般具有持久而强烈的分裂能力,位于根端和茎端较前的部分。初生分生组织是由原分生组织刚衍生的细胞组成,这些细胞在形态上已出现了最初的分化,但细胞仍具有很强的分裂能力,因此,它是一种边分裂、边分化的组织,也可看作是由分生组织向成熟组织过渡的组织。次生分生组织是由成熟组织的细胞,经历生理和形态上的变化,脱离原来的成熟状态(即反分化),重新转变而成的分生组织。如果把二种分类方法对应起来看,则广义的顶端分生组织包括原分生组织和初生分生组织,而侧生分生组织一般讲是属于次生分生组织类型,其中木栓形成层是典型的次生分生组织。
成熟组织 mature tissue
分生组织衍生的大部分细胞,逐渐丧失分裂的能力,进一步生长和分化,形成的其他各种组织,称为成熟组织,有时也称为永久组织(permanent tissue)。各种成熟组织可以具有不同的分化程度,有些组织的细胞与分生组织的差异极小,具有一般的代谢活动,并且也能进行分裂。而另一些组织的细胞则有很大的形态改变,功能专一,并且完全丧失分裂能力。因此,组织的“成熟”或“永久”程度是相对的。而且成熟组织也不是一成不变的,尤其是分化程度较浅的组织,有时能随着植物的发育,进一步特化为另一类组织;相反,有时在一定的条件下,又可以反分化(或脱分化dedifferentiation)成分生组织。成熟组织可以按照功能分为保护组织(protective tissue)、薄壁组织(parenchyma)、机械组织(mechanical tissue)、输导组织(conducting tissue)和分泌结构(secretory structure)。
鞭虫(Trichuris trichura)属刺嘴亚纲毛首目。成虫寄生于人体盲肠内,严重感染时也寄生于阑尾、结肠及直肠等处。虫体前3/5细如鞭状。重度感染出现阵发性腹疼,慢性腹泻及便血等症状
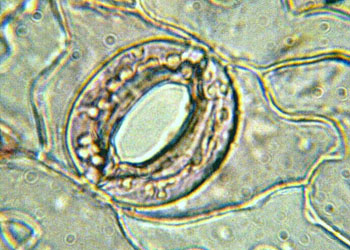
在气生表皮上具有许多气孔(stoma),它们是气体出入植物体的门户。气孔是由二个特殊的细胞即保卫细胞(guard cell)和它们间的开口共同组成的。在有些场合,也有单把开口称气孔的。保卫细胞成肾形或哑铃形,细胞内含有叶绿体,特殊的不均匀增厚的细胞壁,使保卫细胞形状改变时能导致孔口的开放或关闭,从而调节气体的出入和水分的蒸腾。
索虫目(Mermithida)体细长如索,可达50cm。成虫无口囊,头感器16个,由口直接连接咽,肠特化成两行大的营养细胞。幼虫寄生于昆虫及无脊椎动物体内,成虫在土壤或淡水中自由生活
保护组织
(protective tissue)是覆盖于植物体表起保护作用的组织,它的作用是减少体内水分的蒸腾,控制植物与环境的气体交换,防止病虫害侵袭和机械损伤等。保护组织包括表皮(epidermis)和周皮(periderm)。表皮又称表皮层,是幼嫩的根和茎、叶、花、果实等的表面层细胞。它是植物体与外界环境的直接接触层,因此,它的特点与这一特殊位置和生理功能密切有关。表皮一般只有一层细胞,但它不只是由一类细胞组成,通常含有多种不同特征和功能的细胞,其中表皮细胞是最基本的成分,其他细胞分散于表皮细胞之间。表皮细胞呈各种形状的板块状,排列十分紧密,除气孔外,不存在另外的细胞间隙。表皮细胞是生活细胞,细胞一般不具叶绿体,但常有白色体和有色体,细胞内储藏有淀粉粒和其他代谢产物如色素、丹宁、晶体等。茎和叶等植物体气生部分的表皮细胞,外弦向壁往往较厚,并角质化,此外,在壁的表面还沉积一层明显的角质层,使表皮具有高度的不透水性,有效地减少了体内的水分蒸腾,坚硬的角质层对防止病菌的侵入和增加机械支持,也有一定的作用。有些植物(如甘蔗的茎、葡萄、苹果的果实)在角质层外还具有一层蜡质的“霜”,它的作用是使表面不易浸湿,具有防止病菌孢子在体表萌发的作用。在生产实践中,植物体表面层的结构情况,是选育抗病品种,使用农药或除草剂时必须考虑的因素。表皮的结构和角质层纹型,也是植物分类上的一个依据。现在通过电子显微镜的观察研究,对角质层的结构有了进一步的了解,它包括二层,位于外面的一层由角质和蜡质组成,位于里面的一层由角质和纤维素组成。有人提出将这二层合称为角质膜(相当于原来的角质层),而将外层称为角质层,将内层称为角化层。角化层和初生壁之间明显地有果胶层分界。在气生表皮上具有许多气孔(stoma),它们是气体出入植物体的门户。气孔是由二个特殊的细胞即保卫细胞(guard cell)和它们间的开口共同组成的。在有些场合,也有单把开口称气孔的。保卫细胞成肾形或哑铃形,细胞内含有叶绿体,特殊的不均匀增厚的细胞壁,使保卫细胞形状改变时能导致孔口的开放或关闭,从而调节气体的出入和水分的蒸腾。表皮还可以具有各种单细胞或多细胞的毛状附属物。一般认为表皮毛具有保护和防止下面水分丧失的作用。我们用的棉和木棉纤维,都是它们种皮上的表皮毛。有些植物具有分泌功能的表皮毛,可以分泌出芳香油、粘液、树脂、樟脑等物质。根的表皮主要与吸收水分和无机盐有关,因此,它是一种吸收组织(absorptive tissue)。根的表皮细胞具有薄的壁和薄的角质层,许多细胞的外壁向外延伸,形成细长的管状突起――根毛(root hair),从而大大地有利于根的吸收。表皮在植物体上存在的时间,依所在器官是否具有加粗生长而异,具有明显加粗生长的器官,如裸子植物和大部分双子叶植物的根和茎,表皮会因器官的增粗而破坏、脱落,由内方产生的次生保护组织――周皮所取代。在较少或没有次生生长的器官上,例如叶、果实、大部分单子叶植物的根和茎上,表皮可长期存在。周皮是取代表皮的次生保护组织,存在于有加粗生长的根和茎的表面。它由侧生分生组织――木栓形成层形成。木栓形成层平周地分裂,形成径向成行的细胞行列,这些细胞向外分化成木栓(phellem或cork),向内分化成栓内层(phelloderm)。木栓、木栓形成层和栓内层合称周皮。木栓具多层细胞,在横切面中细胞呈长方形,紧密排列成整齐的径向行列,细胞壁较厚,并且强烈栓化,细胞成熟时原生质体死亡解体,细胞腔内通常充满空气。这些特征使木栓具有高度不透水性,并有抗压、隔热、绝缘、质地轻、具弹性、抗有机溶剂和多种化学药品的特性,对植物体起了有效的保护作用。同时也使它在商业上有相当的重要性,可供日用或作轻质绝缘材料和救生设备等。栓皮储、栓皮栎和黄檗(黄柏)是商用木栓的主要来源。栓内层是薄壁的生活细胞,常常只有一层细胞厚,一般只能从它们与外面的木栓细胞排成同一整齐的径向行列,而与皮层薄壁细胞区别开来。在周皮的某些限定部位,其木栓形成层细胞比其他部分更为活跃,向外衍生出一种与木栓细胞不同,并具有发达细胞间隙的组织(补充组织)。它们突破周皮,在树皮表面形成各种形状的小突起,称为皮孔(lenticel)。皮孔是周皮上的通气结构,位于周皮内的生活细胞,能通过它们与外界进行气体交换。
色矛亚纲
(Chromadoria)咽圆柱形,其前后端为球形中间细窄如峡状,咽腺3个,为单细胞腺体。色矛目(Chromadorida):具螺旋形化感器,头感器在极前端,排列成一圈或二圈,口囊内具齿,咽前后端具球形膨大,体表角质层具环状或刻点状装饰。海水、淡水及土壤生活,如色矛虫(Chromadora)。疏毛目(Araeolaimida):头感器排列成3圈,第三圈特化成4个细长头毛,口前端漏斗形,口囊内通常无齿,体表具环纹,但无刻点,主要海产如Plectus。带线虫目(Desmocolecida):身体粗短,具明显的环状,体表装饰有鳞毛、刺、瘤等,身体前端有色素小点或小眼,缺乏明显的口囊,主要海产,淡水及土壤也偶有发现,如链头线虫(Desmoscolex)。单宫目(Monohysterida):头端具分散的刚毛,化感器呈环状,体表光滑或具环纹,有时体表刚毛排列成4或8个纵列。单个卵巢,海水、淡水及土壤均有分布,例如咽管线虫(Siphonolaimus)。
尾感器纲 Phasmida(胞管肾纲 Secernentea)
身体尾端具一对尾感器,无尾腺,化感器不发达,排泄器官为胞管状,位于身体两侧上皮索内。咽腺通常3个,绝大多数为陆生,偶然在淡水中发现,无海产种,分为3个亚纲。
小杆亚纲
(Rhabditida)咽多分为3部分,但末端球内常具有瓣膜。雄性个体具发达的交合囊。小杆目(Rhabditia):小型到中型线虫,头感器均为乳突状,其数目6~12个不等。口囊为长管状,咽可分为3~5部分。卵巢1~2个,在土壤中营自由生活或在脊椎动物体内寄生,例如小杆线虫(Rhabditis)。圆线虫目(Strongylida):口囊为柱形但无区分,雄性具交合囊,是由两个发达的侧叶及一中叶组成,侧叶中有6条放射肋支持,成虫寄生于脊椎动物小肠内,生活史中幼虫期有自由生活的阶段,例如十二指肠钩虫Ancylostoma duodenale(寄生在人小肠内)、美洲板口钩虫Necator americanus、粪类圆线虫Strongyloides stercoralis(生活史包括自生世代和寄生世代,在寄生世代中,成虫主要寄生在 如人、狗、猫等宿主小肠,幼虫可侵入肺、脑、肝、肾等组织器官)。。蛔虫目(Ascaridia):唇片3个或6个,无口囊,咽呈柱形,缺乏肌肉质咽球(蛔虫例外)雄虫有2个等长的交合刺,如人蛲虫Enterobius vermicularia(主要寄生于肠道内,直接感染,儿童感染率极高,夜间爬出肛门产卵,导致肛门瘙痒)、人蛔虫Ascaris lumbricoides(线虫动物代表动物,人体最常见肠道寄生虫之一,感染率高,尤其是儿童)、猪蛔虫Ascaris suum(寄生于猪小肠内,形态和遗传性与人蛔虫极相似)、弓蛔虫Toxocara(寄生于食肉类动物)、禽蛔虫Ascaridia(寄生于鸡等家禽)、副蛔虫Parascaris(寄生于驴、马)、蛇蛔虫Ophidascaris(寄生于蛙、蛇两栖爬行动物)、多宫蛔虫Ascaridia(寄生于爬行动物)。
钩虫成虫寄生于人体小肠的上段,前端具口囊、唇片退化,口囊内具有成对的钩齿,具切割作用
钩虫成虫在小肠内寄生时,咬破肠粘膜,吸食血液,同时虫体可分泌抗凝血酶,使伤口处不停的渗血,使病人严重贫血
成虫在小肠内交配并产卵,每头雌虫日产卵数万粒,卵随寄主粪便排到体外,卵在适宜环境中可孵化为杆状幼虫
美洲(板口)钩虫Necator americanus与十二指肠钩虫在形态上相似,但体较小,口囊内无钩齿,腹侧有一对半月形的见板,雄性毛端交合伞其背肋小枝有2个分叉。属热带型,主要分布于热带、亚热带地区,我国南方多于北方
十二指肠钩虫成虫在小肠内交配并产卵,每头雌虫日产卵数万粒,卵随寄主粪便排到体外,当温度适宜时,卵在松软的土壤中1~2天即可孵化为一期
杆状幼虫
,幼虫具细长口腔,虫体300μm左右,经2~3天后脱皮成二期杆状幼虫
二期杆状幼虫在土壤中发育脱皮成
丝状幼虫
,即感染期幼虫(
感染蚴
)。感染期幼虫具有群集习性,在土壤中可存活三个月左右。当人赤足在土壤上行走,或手接触土壤时,感染蚴会从手指之间或足趾间薄嫩皮肤处进入皮内,再随血液或淋巴液移行,经右心、肺,然后再逆行至咽,经吞咽进入小肠,在小肠内发育成成虫
钩虫
成虫寄生于人体小肠的上段,人体寄生的钩虫主要有两种,即十二指肠钩虫(Ancylostoma duodenale)及美洲钩虫(Necator americanus),我国北方以前者为主,南方多感染美洲钩虫。成虫前端具口囊、唇片退化,口囊内具有成对的钩齿,具切割作用,雄虫尾端具交合囊(Copulatory bursa)及刺。成虫在小肠内交配并产卵,每头雌虫日产卵数万粒,卵随寄主粪便排到体外,当温度适宜时,卵在松软的土壤中1~2天即可孵化为一期杆状幼虫,幼虫具细长口腔,虫体300μm左右。经2~3天后脱皮成二期杆状幼虫,在土壤中发育5~8天后再脱皮成丝状幼虫,即感染期幼虫。感染期幼虫具有群集习性,在土壤中可存活三个月左右。当人赤足在土壤上行走,或用手接触土壤时,感染期幼虫会从手指之间或足趾间薄嫩皮肤处进入皮内,再随血液或淋巴液移行,经右心、肺,然后再逆行至咽,经吞咽进入小肠。在小肠内再脱皮二次,发育成成虫。感染期幼虫从皮肤进入人体到雌虫产卵,约需5~7周,十二指肠钩虫在人体内可存活1~7年,美洲钩虫存活更长。幼虫在侵入寄主皮肤后,可刺激皮肤出现丘疹、皮炎等,但数日后可消失。幼虫在人体内移行时,引起寄主咳嗽、发烧、咳痰、哮喘等症状,症状的轻重与侵入体内虫体的数量相关。成虫在小肠内寄生时,咬破肠粘膜,吸食血液。同时虫体可分泌抗凝血酶,使伤口处不停的渗血。造成肠壁严重机械损伤。成虫有不断更换咬吸部位的习性,因此造成新老伤口同时流血不止,寄主大量新鲜血液由肠壁伤口处流失,使病人严重贫血,病人出现头晕眼花、心跳气短、苍白无力,甚至浮肿、贫血性心脏病,严重丧失劳动能力。一些病人还出现“异嗜症”,即喜食生米、泥土、纸张等非正常食品,据研究这是由于严重贫血引起的缺铁症,如单独补充铁剂,异嗜症状可缓解。我国黄河以南地区农村的感染率可达人口的20%左右,个别地区可高达35%。
人蛔虫是人体感染最普遍的一种寄生线虫,寄生于人体小肠内
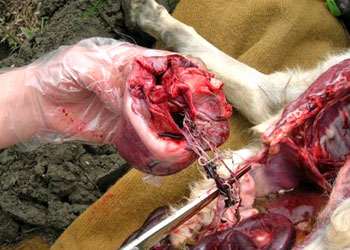
体内大量蛔虫成虫寄生,会出现成虫扭曲成团造成肠梗阻
从一5岁男孩坏死肠道中取出的蛔虫,近2千克重
蛔虫成虫在人体小肠内交配并产卵,每头雌虫日产卵量可高达20万粒,卵随寄主粪便排到体外。卵在适宜环境中发育成具感染能力的卵(胚胎卵)
蛔虫卵感染无需中间寄主,经人误食可直接进入小肠,进而孵化出幼虫,幼虫穿过肠粘膜进入静脉,并随血液在体内循环,经过肝、心脏,最后到达肺部
蛔虫幼虫在肺泡内寄生,在肺泡内脱皮两次,随咳嗽等动作沿气管逆行又回到咽部,再经吞咽动作又进入消化道中,进入小肠后再脱皮一次,数周后发育成成虫
人蛔虫
(Ascaris lumbricoides)是人体感染最普遍的一种寄生线虫,寄生于人体小肠内,属于大型的人体寄生线虫。雌虫长达20~35cm,雄虫15~30cm,由卵进行传播,成虫与幼虫均在人体内寄生。成虫在人体小肠内交配并产卵,每头雌虫日产卵量可高达20万粒,卵随寄主粪便排到体外。卵到外界在20~30℃、阳光充足、潮湿松软的土壤中经二周后在310卵内发育成幼虫,一周后幼虫在卵内脱皮一次成为具感染能力的卵。新排出的卵没有感染力。人如吞食了感染卵,卵到小肠后则幼虫孵化,幼虫穿过肠粘膜进入静脉,并随血液在体内循环,经过肝、心脏,最后到达肺部,幼虫在肺泡内寄生,在肺泡内脱皮两次,随咳嗽等动作沿气管逆行又回到咽部,再经吞咽动作又进入消化道中,进入小肠后再脱皮一次,数周后发育成成虫,人体自虫卵感染到雌虫产卵,约需60~70天,成虫在人体内存活一年左右。人如少量感染蛔虫,并不引起明显症状,如果严重感染则对人体造成很大危害。幼虫在人体内移行时,释放出免疫原性物质,引起寄主局部或全身的变态反应,如肺部炎症、痉挛性咳嗽、体温上升等。成虫在小肠内寄生,引起小肠粘膜机械性损伤,以致消化吸收不良,病人腹疼、食欲不振,严重时儿童会出现贫血、发育障碍等症状。体内大量成虫寄生,会出现成虫扭曲成团造成肠梗阻或成虫侵入胆囊,造成胆囊炎、胆道穿孔、胰腺炎、腹膜炎等。蛔虫是世界性分布的人体寄生虫,在我国感染很普遍,特别是在农村。蛔虫成虫产卵量大、虫卵有很强的理化抗性,直接感染不需要中间寄主,人们生食蔬菜及不卫生的生活习惯等都造成了蛔虫的广泛传播。因此积极治疗病人、管理粪便、改善环境条件及注意个人卫生是控制蛔虫流行的重要手段。
蛲虫寄生于人的盲肠、结肠、直肠等部,雌蛲虫在午夜时爬到寄主肛门处产卵,致使肛门奇痒,影响睡眠
当患者由于雌虫及卵的刺激用手搔抓肛门时,虫卵可经污染的手指进行自体感染,亦可经衣被、患者用具,甚至空气进行传播
蛲虫进入消化道后,前端钻入肠粘膜,吸取营养。患者多为儿童,严重时会影响睡眠、食欲不振、烦躁、消瘦等症状出现
蛲虫
(Enterobius vermicularis)是寄生于人的盲肠、小肠下段的一种小型线虫,又名蠕形住肠线虫,雌性成虫体长8~13mm,雄虫2~5mm。雌、雄头端均有角质膨大形成的翼。成虫在寄生部位交配,交配后雄虫死去,雌虫子宫内充满卵粒后向下移行。夜间寄主入睡后,雌虫到寄主肛门处产卵,产卵后雌虫多数死亡。偶有雌虫仍爬回直肠。虫卵在外界温度适宜,氧气充足的条件下,经数小时后即变成具感染能力的卵。当患者由于雌虫及卵的刺激用手搔抓肛门时,虫卵可经污染的手指进行自体感染,亦可经衣被、患者用具,甚至空气进行传播,也偶有虫卵在肛门外孵化,然后幼虫再爬回直肠。具感染力的虫卵进入人体后在小肠内孵化为幼虫,幼虫沿小肠下行时脱皮两层,至结肠再脱皮一次发育为成虫。自感染到雌虫产卵约需一个半月左右,成虫寿命2~4周。蛲虫患者多为儿童,特别是在儿童集体生活的条件下易于传播流行。蛲虫也是世界性分布的寄生虫。患者轻度感染时症状不明显,严重感染会影响睡眠、食欲不振、烦躁、消瘦等症状出现。治疗病人、注意个人卫生、加强家庭及托儿机构的卫生是控制其流行的手段。
旋尾亚纲
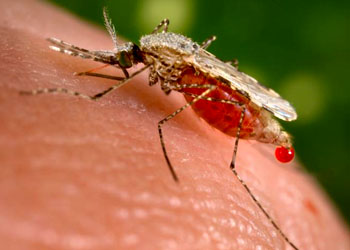
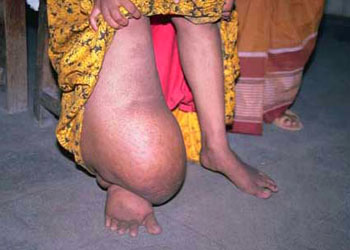
(Spiruria)口囊呈球状或圆柱状或不明显,咽前端窄、后端膨大,其中绝无瓣膜。雄虫的交合刺不等长。旋尾目(Spiruria):身体细长,中等大小,尾部盘曲,故名旋尾虫,头部具两个侧唇(或4个或更多的唇),口囊角质化,或具齿,交合刺不等长。成虫寄生于脊椎动物的消化道及呼吸道中,中间寄主为节肢动物,如棘颚口线虫(Gnathostoma),寄生于猫、犬体内;斑氏丝虫Wuchereria bancrofti,寄生于人体淋巴系统,可引起组织增生,使下肢、阴囊等处畸形,形成“橡皮病”。
班氏丝虫(Wuchereria bancrofti)多寄生于人体深部淋巴系统中,如下肢、阴囊、腹股沟等部位,雌雄成虫需交配生殖,卵胎生,没有自由生活的幼虫阶段,由中间寄主蚊子进行传播
在人体内丝虫的雌虫可直接产出
微丝蚴
,微丝蚴进入血液,白天停留在肺血管中,夜间人睡眠后,微丝蚴出现于人外周末稍微血管中,微丝蚴在人体内可存活2~3个月
微丝蚴可随蚊的叮咬进入
蚊
胃中,在蚊的肌肉中发育成感染期幼虫,再迁移至蚊体腔,最后到唾腺中,当蚊虫再度吸血时,将感染期幼虫传入人体,最后在淋巴管或淋巴结内发育为成虫
丝虫反复感染及淋巴管炎症周期性发作,导致淋巴组织不断增生肿大,皮脂腺及汗腺萎缩而出现“橡皮肿”,班氏丝虫患者出现睾丸急性炎症及阴囊橡皮肿,个别病人阴囊可达10~20公斤
马来丝虫患者多出现下肢橡皮肿,病人丧失劳动力,但患者病程较长,最长者可达45年之久。治疗病人、消灭蚊子及其孳生地、防止被蚊虫叮咬是控制丝虫流行的有效途径
犬心丝虫(Dirofilaria Immitis)寄生于犬、猫的右心室及肺动脉,造成严重的循环不良及肺功能障碍,并使肝、肾等重要器官严重受损,最后导致寄主死亡,
丝虫
是寄生于人体淋巴系统中的一种线虫,寄生于人体的丝虫共有8种,我国只有两种,即班氏丝虫(Wuchereria bancrofti)及马来丝虫(Brugia malayi)。前者多寄生于人体深部淋巴系统中,如下肢、阴囊、腹股沟等部位,后者多寄生于下肢浅部淋巴管中,两者均由中间寄主蚊子进行传播。班氏丝虫成虫体长雌虫75~100×0.2~0.3mm;雄虫30~45×0.1~0.15mm,马来丝虫略小,雌虫体长约55mm,雄虫约25mm左右。雌雄成虫需交配生殖,卵胎生,没有自由生活的幼虫阶段。在人体内丝虫的雌虫可直接产出微丝蚴,微丝蚴进入血液,白天停留在肺血管中,夜间人睡眠后,微丝蚴出现于人外周末稍微血管中。微丝蚴在人体内可存活2~3个月。夜间当蚊子叮咬患者时,微丝蚴进入蚊胃中,随后进入蚊的肌肉中发育,在蚊体内脱皮两次。班氏丝虫的幼虫在蚊体内经过二周,马来丝虫的幼虫在蚊体内经一周左右可发育成感染期幼虫,这时感染期幼虫离开蚊的肌肉进入蚊体腔,再到唾腺中,当蚊虫再度吸血时,将感染期幼虫传入人体,最后在淋巴管或淋巴结内发育为成虫,一般人感染3个月之后,在血液中即可查到微丝蚴。成虫在人体内可存活十几年。人被丝虫感染后,初期出现皮肤过敏及炎症反应,主要危害由成虫引起,由于成虫在淋巴管及淋巴结内的机械刺激作用及虫体代谢或分解的毒素,引起局部的淋巴系统发炎,由肿疼处离心方向出现红线,俗称“流火”。这时病人寒高烧、头疼等症状,病程持续一周左右自行消失。由于反复感染及淋巴管炎症周期性发作,每年发作数次,致使淋巴管壁组织不断增生,以致慢性堵塞,使阻塞的淋巴管下端的淋巴液不能回流,刺激皮肤及皮下组织增生,使之变粗变厚,皮脂腺及汗腺萎缩而出现“橡皮肿”,通常还伴随有皮肤溃疡。班氏丝虫患者出现睾丸急性炎症及阴囊橡皮肿,个别病人阴囊可达10~20公斤。马来丝虫患者多出现下肢橡皮肿,病人丧失劳动力,但患者病程较长,最长者可达45年之久。我国江南流行区感染率可高达20%左右。治疗病人、消灭蚊子及其孳生地、防止被蚊虫叮咬是控制丝虫流行的有效途径。
秀丽小杆线虫(Caenorhabditis elegans)自由生活于土壤中,是发育生物学和神经生物学的研究的模式生物,如对线虫细胞谱系、细胞程序性死亡和小分子RNA的研究等
粪类圆线虫Strongyloides stercoralis生活史包括自生世代和寄生世代,在寄生世代中,成虫主要寄生在 如人、狗、猫等宿主小肠,幼虫可侵入肺、脑、肝、肾等组织器官,引起粪类圆线虫病
小麦线虫(Anguina tritica)从幼虫到成虫寄生在小麦植株上,并在麦穗上形成虫瘿,在其中产卵孵化,每个虫瘿中可有数千至数万条幼虫,虫瘿随麦粒播入土中,幼虫出来再感染新的小麦
双胃线虫亚纲
(Diplogasteria)多数为小型线虫,体长3~4mm,体表具环纹,有刻点,唇不发达,咽可分为4部分,有瓣膜,但一定在峡之前,此点不同于其他两亚纲。双胃线虫目(Diplogasterida):口囊变化很大,多数具齿,咽的特征如亚纲所述,多在昆虫体内寄生,如双胃线虫(Diplogaster)。垫刃线虫目(Tylenchida):唇区光滑发达,化感器位于唇上,口囊内具长刺,咽内瓣膜位于峡之前,排泄器官仅有一条纵管,分布于一侧。雌虫具1~2个卵巢,雄虫有一对尾感器,寄生于昆虫及高等植物体内,如垫刃线虫(Tylenchulus);小麦线虫(Anguina tritici)寄生于小麦子房上,使麦粒形成虫瘿。
线虫动物门的形态与生理
寄生种类的线虫具发达的
角质层
,是上皮细胞的分泌物,它限制了身体的生长,所以线虫在生长过程中经过数次到多次的
脱皮
现象。角质层下为合胞体的表皮层,肌肉层只有较厚的纵肌层,而无环肌层,因此线虫只能摆动,不能伸缩运动
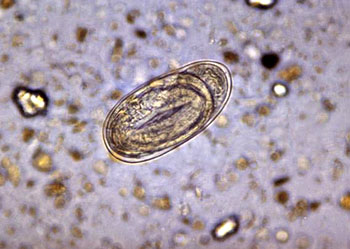
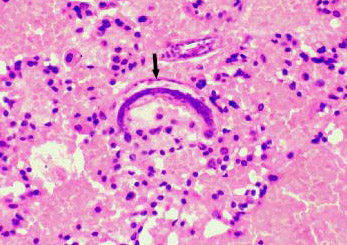
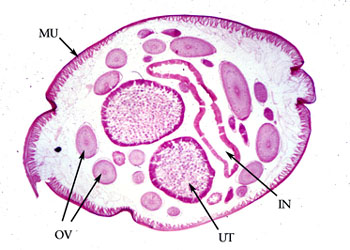
雌性个体常有两个卵巢,少数个体仅一个卵巢,卵巢管长短不等,相对排列或平行排列,卵巢后端为输卵管、子宫。子宫的上端为受精囊,用以贮存交配后的精子(图为雌性蛔虫的横切面,UT为子宫,OV为卵巢)
线虫主要为雌雄异体及异形,雄性个体较小,后端弯曲成钩状。生殖腺均为管状,生殖细胞是由生殖腺中一个大的末端细胞(terminal cell)发生,在其通过生殖腺的过程中,生长成熟(图为人蛔虫,上雌下雄)
外形
:绝大多数自由生活的线虫是小型动物,体长一般不超过2.5mm,多数在1mm左右,陆生的大型个体可长达7mm,海产的可达5cm。但寄生种类最长的可达1m,例如寄生于猪体内的膨结线虫(Dioctophyma)。身体多圆柱形、两端略尖细,身体没有分区,例如小杆线虫(Rhabditis)。身体前端有口、口周围有唇(lip),原始的种类具有6个唇片,如Plectus,其他种常有3个唇片,唇上及其周围有感觉乳突(papillae)或感觉毛(bristles)等感觉器官。唇及感觉器的数目、排列及形态是线虫分类的重要依据之一,它们多呈放射状排列,有人认为这种对称形式说明线虫的祖先是固着生活的,是以尾端粘液腺分泌粘液附着在底部,以后垂直延长身体,脱离围着生活,而进化成现代的线虫。线虫体表光滑或由角质层特化形成环、嵴、瘤及纵线等。使体表呈现出各种装饰,身体近后端腹面有肛门,在身体中段、腹中线上雌性个体有生殖孔。
体壁及运动
:线虫的体壁亦由角质层、上皮细胞及肌肉组成。线虫的角质层结构较复杂,可分为三层,其最外层为一层鞣化蛋白质,表现出环纹;中层为均质的,内层为胶原蛋白构成的支持柱层。寄生的蛔虫类等,其内层由于纤维排列的方向不同,又可分为三层,因此使角质层表现出一定的弹性。角质层是上皮细胞的分泌物,它限制了身体的生长,因此线虫在生长过程中经过数次到多次的脱皮现象。在脱皮前,角质层中有用的物质被虫体吸收,然后上皮细胞重新分泌新的表皮,使旧表皮与上皮细胞分离,并最后完全脱落,使虫体得以生长。研究指明,线虫的脱皮是由神经环上的神经细胞分泌出激素(hormone),激素促使排泄细胞分泌脱皮液,脱皮液可溶解旧表皮而造成脱皮,脱皮现象仅出现在幼虫期,成虫仅能增加角质层的厚度而不再脱皮。角质层内为上皮细胞,上皮细胞或界线清楚或为合胞体。在身体背、腹中线及两侧、上皮细胞向内凸出形成4条纵行的上皮索。上皮细胞核仅局限在索中,并排列成行。这4条上皮索在体表明显可见,分别称为背线、腹线及侧线。上皮之内为肌肉层,线虫缺乏环肌,只有纵肌,分布在上皮索之间,肌肉为斜纹肌。肌细胞的基部为可收缩的肌纤维,端部为不能收缩的细胞体部,它的功能可能是贮存糖元,核位于细胞体部。细胞体部的原生质延伸形成线状,分别连接到背索与腹索内的神经上,在那里接受神经支配。其他动物是由神经发出分支分布到肌肉上进行支配,而不是像线虫由肌肉延伸到神经处,去接受支配。体壁之内为假体腔,其中充满体腔液,体腔液内没有游离的细胞,但有体腔细胞固着在肠壁及体壁上,体腔液除了担任输送营养物及代谢物之外,还有抗衡肌肉收缩所产生的压力,起着骨骼的作用。线虫的运动是由纵肌的收缩及角质层的弹性改变而共同完成。当背纵肌收缩时,腹面角质层中的纤维拉长,当背纵肌松弛时,腹面角质层中的纤维恢复,因此表现出身体背腹方向的蛇行运动。一些种类体表具刺、环等,可做短距离的爬行或游泳运动。
取食与消化
:线虫的食性很广泛,许多自由生活的线虫是肉食性的,以小形的动物为食,也有许多种为植食性的,以藻类及植物根部细胞及其内含物为食,还有的种类是细菌及沉积物的取食者,是以溶解的动、植物尸体或有机颗粒为食,它们构成了数量最大、分布最广的细菌及真菌的取食者,在生物链中起着重要作用。线虫的消化系统结构简单,为一直形管,前端口后为一管状或囊状的口囊(buccal capsule),口囊内壁角质层加厚,形成不同形状或不同数目的嵴、板、齿等结构,用以切割食物。特别在肉食性的种类较发达。有的种类在口囊中形成一中空或实心的刺,用以穿刺食物或抽吸食物汁液。口囊之后为咽,咽常形成一个或几个咽球,由于肌肉细胞的加厚,咽腔在断面上呈三放形,三放中的一放总是指向腹中线,构成线虫咽的一个特征。咽的周围有成对的咽腺,可分泌消化液,咽腺可开口在咽前端。咽由于很厚的肌肉层,具有泵的作用,可由口抽吸食物进入咽及肠。咽后紧接为中肠,是由单层上皮细胞组成,中肠的两端均有瓣膜,以阻止肠内食物逆流。中肠后为短的直肠,最后以肛门开口在近末端的腹中线上。线虫的咽腺及中肠的腺细胞产生消化酶,在中肠内进行食物的消化,并在肠壁细胞内完成细胞内的消化。线虫类或通过体表进行气体交换或行低氧呼吸,它没有专门的呼吸及循环结构。
排泄及水分调节
:线虫的排泄器官属于原肾型,而结构不典型,是由腺型细胞或由细胞形成的管完成排泄功能。原肾型特点是由外胚层内陷形成;只在体表有开口,体内没有开口。但这些动物的代谢废物更多的是从体表排出,因此原肾排泄系统的主要功能是保持体液的稳定,保持动物体内的水盐平衡(这与原生动物伸缩泡的作用类似)。原始的种类只有1~2个大型的腺细胞进行排泄及水分调节,这种腺细胞具有长颈,位于咽的周围,也称肾细胞(renette cell),联合开口在神经环附近的腹中线上,如小杆线虫。另一些种,腺型肾细胞延伸成管型排泄器,多呈H型管,两个侧管在前端以一横管相连分别位于侧上皮索内,最后经过共同的小管以排泄孔开口在前端腹中线上,例如驼形线虫(Camallanus)。蛔虫(Ascaris)的排泄管也呈H形,只是横行管成网状,侧管前端不发达,而且是由一个腺细胞特化形成。线虫的腺细胞及排泄管中没有鞭毛或纤毛,不呈焰细胞状。线虫的代谢产物主要是氨,可以通过体壁及消化道而排出。线虫的排泄器官对维持其体腔液的压力是十分重要的,水可以通过口及体壁进入体内,过多的水分可通过排泄器官排出。实验证明:海产的种类可以在氯化钠的低渗液中进行调节,而不能在高渗液中调节;淡水及陆生的种类相反,维持体腔液低渗于周围环境;陆生线虫还可以在高度干燥条件下,像轮虫一样通过失水,降低代谢速率,处于隐生状态,来渡过干燥可长达数年之久。
神经与感官
:线虫的神经结构与其他假体腔动物大致相似,在咽的周围有一环状的脑,环的两侧膨大成神经节,由脑环向前后各分出六条神经,前端的神经分布到唇、乳突及化感器等。向后的六条神经中,一条为背神经,一条为腹神经,二对侧神经,两对侧神经离开脑环后很快合并成一对,最后的这四条神经分别位于相应的纵行上皮索内,其中腹神经最发达,由腹神经发出分支到肠及肛门。腹神经索中包括运动神经纤维及感觉神经纤维,背神经索中主要为运动神经纤维,侧神经索中主要为感觉神经纤维。另外在脑环周围,神经细胞集中,形成神经节状。线虫的感觉器官主要分布在头部及尾部两端。头部包括唇、乳突、感觉毛,统称为头感器;还有一个特殊的化感器(amphid)。唇及乳突是头部的角质突起,有脑环发出的神经进行支配。感觉毛在头部较发达是一种触觉感受器,实际上是一种改变了的纤毛,有的种感觉毛周围还有腺细胞围绕。化感器是线虫特有的一种感觉器,位于身体前端一侧,它是体表的一个内陷物,呈囊状、管状、螺旋状等各种形态,成为线虫分目的依据之一,其内端为盲端,外端开口,它是一种化学感受器,也常有腺细胞伴随。电子显微镜的研究已经证明化感器的感觉突实际上也是改变了的纤毛,过去一直认为线虫类不存在任何纤毛,而化感器是改变了的纤毛,这种特征便将线虫和其他有纤毛的动物在进化上联系起来。在水生种类特别是海产种类化感器发达,而陆生及寄生种类则退化了。水生种类在咽的两侧还有一对眼点,是视觉器官其中色素细胞分散或排列成杯状。线虫的尾端也有一对单细胞腺体称尾感器(phasmid),分别开口在尾端两侧,它也是一种腺状感受器,这种感受器在寄生的种类发达。雄性个体尾端交配器周围也有感觉乳突及感觉毛。
生殖与发育
:线虫主要为雌雄异体及异形,雄性个体较小,后端弯曲成钩状。生殖腺均为管状,生殖细胞是由生殖腺中一个大的末端细胞(terminal cell)发生,在其通过生殖腺的过程中,生长成熟。少数线虫生殖细胞来自整个生殖腺的管壁细胞。雄性生殖系统通常具有一个精巢,少数种是两个。后端连接输精管及其膨大的贮精囊,向后为肌肉发达的射精管(ejaculatory duct),开口到直肠或泄殖腔。射精管周围有数目不等的前列腺(prostatic gland)。大多数线虫雄性泄殖腔向外伸出两个囊,每个囊中有一角质的交合刺(spicule),有肌肉牵引可自由的由泄殖孔伸出体外交配时用以撑开阴门。交合刺的长短、形态因种而异,也是分类标准之一。还有的线虫交合刺的背壁有角质小骨片,愈合成副刺(gubernaculum),以控制交合刺的运动。线虫的精子具鞭毛或不具鞭毛而呈囊状、球状等。雌性个体通常有两个卵巢,少数个体仅一个卵巢,卵巢管长短不等,相对排列或平行排列,卵巢后端为输卵管、子宫。子宫的上端为受精囊,用以贮存交配后的精子,两个子宫后端联合,经肌肉质阴道,以雌性生殖孔开孔在身体近中部腹中线上。线虫需交配受精,交配时雄虫以尾端对准雌性生殖孔,再以交合刺撑开阴门,由交合囊及射精管肌肉的收缩送精子入雌体,精子经鞭毛运动或变形运动到达子宫上端,在此与卵融合,卵受精后形成厚的受精膜,变硬后形成卵的内壳,卵沿子宫下行时,子宫的分泌物形成卵的外壳,卵壳外层的形状常作为寄生线虫分类的依据。卵常在子宫内时已开始发育。极少数的线虫是雌雄同体的,如小杆线虫目的一些种,并行自体受精。也有极少数陆生线虫可行孤雌生殖。自由生活的线虫产卵量较少,海产种类一般仅产数十粒卵,陆生种产卵较多,可达数百粒。寄生种类产卵量极大,每日可产卵数千到数十万粒。线虫的卵是决定型卵,分裂球排列不对称,许多种细胞分裂在胚胎期已经完成,除了生殖系统之外,其他的器官细胞数已固定,孵化后幼虫的生长是通过细胞体积的增加而实现。幼虫期一般脱皮4次,前两次脱皮常在卵壳中进行,成年后不再脱皮。
名称
生殖方式
寄生部位
感染方式
感染虫态
主要危害
蛔虫
卵生
小肠
经口感染
人食入感染性虫卵
夺取营养或引起胆道蛔虫病、肠穿孔
钩虫
卵生
小肠
皮肤接触
丝状幼虫通过足等处皮肤进入人体
贫血、水肿,影响儿童发育
丝虫
卵胎生
成虫在人体淋巴组织中,幼虫在微血管中
蚊吸血传播
丝状幼虫随蚊吸血进入人体
淋巴管炎症、头疼、疲倦或引发四肢、阴囊等形成“象皮肿病”
蛲虫
卵生
大肠
经口感染
人食入感染性虫卵
肛门、会阴等处奇痒,睡眠不安,严重时影响儿童发育
鞭虫
卵生
盲肠
经口感染
人食入感染性虫卵
轻者无症状,严重时有腹痛、恶心、呕吐等现象