海绵动物门
以具碳酸钙构成的骨针为主要特征。全部海产,大多生活在沿岸浅海。三种沟系均存在,型一般很小,常呈暗灰色或灰色,一般在10cm之下。骨针顶端常突出体外,致使海绵呈刺状;典型的钙质海绵为辐射对称的瓶状,顶端是出水口,常被竖立的长单轴骨针围绕,称出水口Z。骨针大小不一,但在分类学上都算作大骨针,有3种基本型:单轴骨针、三辐骨针和四辐骨针,形状多种多样。分两个目:同腔目(Homocoela):体呈单沟型,体壁薄,无褶叠,领细胞连续分布于中央腔,例如白枝海绵(Leucosolenia)。异腔目(Heterocoela):体双沟型或复向型,体壁较厚、褶叠,领细胞分布于放射管或鞭毛室内,例如毛壶(Grantia)。
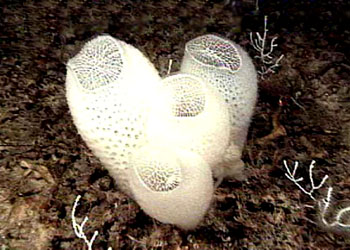
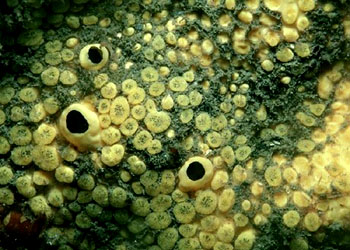
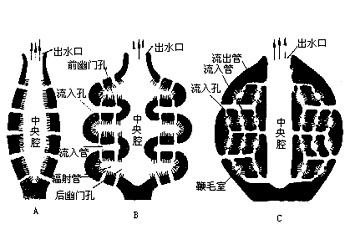
海绵动物都是固着生活的,它们的形状、大小、色泽在不同的种类差别很大。少数种类单体生活,例如毛壶(Grantia),身体呈放射对称、呈柱状或瓶状,体长仅数毫米到一厘米。又如佛子介(Hyalonema)等,身体呈杯状或筒状,它们以身体的基部固着,或以基部伸出的骨针束固着在海底。绝大多数海绵动物为群体生活,群体中的个体有的界线明显,例如群体白枝海绵(Leucosolenia),有的种群体中的个体界线不明显,如淡水海绵(Spongilla),它们往往没有固定的形状,整个的群体常受附着的基底、空间、水流等环境的因素所影响,例如附着在岩石表面的常呈片状,附着在柱上的群体呈筒状,附着在岩缝中的群体常成簇状,即使相同的种也常因附着的基底不同而形成不同形状的群体。群体的体积一般较大,最大的群体直径可达1m、高2m。许多种海绵身体呈明亮的颜色,如桔红色、黄色、绿色、紫色、褐色等。少数种类呈灰色、白色。其中绿色的种类往往是由于体内共生的藻类所致,其他色彩均由体内的色素所形成,其体色出现的意义尚不清楚,可能具有警戒及保护的作用。海绵动物身体的基本结构是由两层细胞围绕中央的一个空腔所组成。游离的一端有一个大的出水口(osculum)使中央腔(central cavity)与外界相通。构成海绵动物体壁的两层细胞在不同的种类组成复杂程度不同的沟系,根据沟系可以将海绵动物的身体结构分为三种类型。
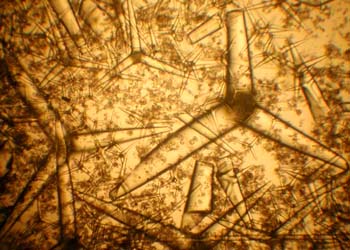
海绵动物除了个别的科没有骨骼之外,其他所有的种类都是具有骨骼的,骨骼是海绵动物的一个典型特征,是用以分类的重要依据之一。海绵动物的骨骼有骨针(spicule)及海绵丝(sponginfiber)两种类型,它们或散布在中胶层内,或突出到体表,或构成网架状。骨骼具有支持及保护身体的功能。骨针的成分或是由碳酸钙组成钙质骨针,或是硅质骨针,其中还都可能包括微量的铜、镁、锌等离子。骨针按其大小又可分为大骨针(megasclere),构成支持身体的骨架;及小骨针(microsclere),它散布在中胶层内,以支持体壁中的管道部分。小骨针仅存在于硅质海绵中。从形态上骨针可以分为多种,其中常见的有(1)单轴骨针(monaxons),即沿一个轴生长形成的骨针,轴或直或弯,轴的两端或相似或不相似,末端或尖或具有其他改变;(2)四轴骨针(tetraxons),也称四放骨针(quadriradiate),这种骨针在一个平面上有四个放射端,但常因丢失一些放射端而变成三放、二放或一放型,三放骨针是钙质海绵纲动物中最普通的一种骨骼;(3)三轴骨针(triaxons),它的三个轴相互以直角愈合,因而呈六放型(hexactinal),这种也常减少末端而改变放数,其末端可以弯曲、分枝、或具钧、具结等变化而形成了多种形态;(4)多轴骨针(polyaxons),由中心向外伸出多射,形成星壮,这种类型多见于小骨针。不同种的海绵,各种骨针或彼此分离,或按一定结构形成疏松的或坚实的网架以支持身体,因此可根据骨针的类型、数量及排列而作为海绵动物分类的依据。海绵丝是一种纤维状骨骼,它是由硬蛋白(scleroprotein)组成,它们或单独的存在于海绵动物体壁内,或与硅质骨针同时存在。许多小的硅质骨针埋在海绵丝中,形成有效地支持物。许多大型群体海绵常同时存在着这两种骨骼。海绵动物的骨针及海绵丝都是由中胶层中的变形细胞特化形成的造骨细胞所形成。单轴的钙质骨针是由一个造骨细胞分泌形成,骨针形成时,造骨细胞核先分裂,并在双核细胞的中心出现一个有机质的细丝,然后围绕这一细丝沉积碳酸钙,随着骨针的逐渐增长,双核细胞也分成两个细胞,并分别加长骨针的两端,最后形成一个单轴骨针。同样,三轴骨针是由三个造骨细胞聚集在一起,每个细胞也随着有机质细丝的形成而分裂一次,形成六个细胞,碳酸钙围绕有机质细丝沉积愈合的结果形成了一个三轴型骨针。海绵丝是由许多造骨细胞联合形成,先是由少数细胞形成分离的小段,然后再愈合成长的海绵丝。在寻常海绵纲动物中,这些海绵丝再相互联结形成网状骨架。
海绵动物是固着生活的,没有移位的运动,仅能通过体表扁平细胞和孔细胞的收缩而略微改变身体的体积。许多双沟型及复沟型海绵,在进水小孔及出水口的四周扁平细胞特化形成类肌细胞(myocyte),类肌细胞的形状及收缩性能有些类似平滑肌,它的收缩可以调节水流出入的速度,当在恶劣环境时,例如污水、暴露于空气中等,类肌细胞可以关闭小孔或出水口,环境改善后,类肌细胞松弛,小孔又重新开放,这种反应是极为缓慢的,有时数分钟后才能看到微小的变化。海绵动物的生理活动是依靠于通过身体的水流而进行,水流可以带进食物及氧气、带走代谢废物、生殖细胞,总之靠水流完成生长、发育及繁殖。有人观察过一种白海绵(Leucandra),这是一种复沟系小型的钙质海绵,流经出水口的水流速度是8.5cm/s,据估计直径1cm、高10cm的白海绵大约有225万个鞭毛室,每天有22.5L的海水流过身体。出水口狭窄,水流速度很快,而鞭毛室数量极多,其内水流缓慢,这样便于领细胞有更多的时间从水中摄取食物及气体交换。水在体内的流动是由领细胞的鞭毛打动所引起,鞭毛由基部向端部旋转运动,使水流进入鞭毛室。同一鞭毛室的鞭毛运动既不同步,也不互相协调,但鞭毛的方向都是指向后幽门孔。许多复沟型海绵在鞭毛室的出口处有一中央细胞,它的收缩可变动位置以调节水流的流量,甚至可以完全关闭后幽门孔而阻止水由鞭毛室流出。海绵动物取食各种有机物颗粒,食物的选择主要是看有机物颗粒是否能进入入水孔。有人用几种海绵动物做实验,结果发现食物中80%是细小的有机质颗粒,20%为细菌、鞭毛虫类及其他极小的浮游生物。食物随水流进入鞭毛室,水流速度在此减慢,有利于领细胞的捕食,领细胞领部的微绒毛可以粘着微小的食物颗粒,然后吞噬,较大的食物颗粒也可以被入水小管的扁平细胞所吞噬。食物进入领细胞后它可进行部分的细胞内消化,部分食物转移到中胶层的变形细胞,在变形细胞中进行食物的消化。消化后的营养物或贮存在变形细胞内,或转移到其他细胞,不能消化的食物残渣仍由变形细胞运出。海绵动物没有专营呼吸与排泄的细胞,而是当水流过身体时,大多数细胞均可与水接触,各自独立完成呼吸与排泄的机能。许多淡水海绵,大多数细胞内具有一到几个伸缩泡,这些伸缩泡像原生动物一样,担任着调节水与盐份的平衡。海绵动物没有神经结构,对刺激的反应常是局部的、缓慢的,对刺激反应的大小是依赖于刺激的强弱。信息物质的传递是通过中胶质中的扩散作用、游离变形细胞及固定细胞彼此的接触而进行。尚未发现海绵动物具有电传导。