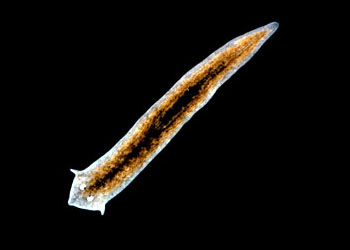
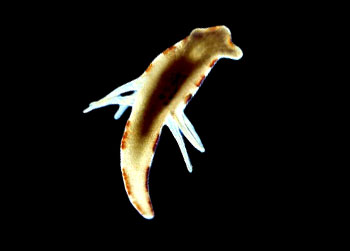
扁形动物门
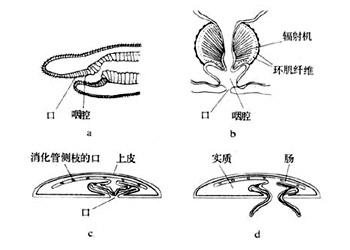
扁形动物的排泄系统为原肾型(protonephridium type),主要由排泄管、毛细胞和焰细胞(flamecell)组成,起源于外胚层的内陷,主要功能是调节体内水分渗透压,同时也排出一些废物
扁形动物出现了原始的梯形神经系统,神经细胞向前集中形成脑及脑向后发出若干纵神经索,在纵轴之间有横神经连接为梯形