涡虫纲
吸虫纲
绦虫纲
你的位置:
首页
>
动物界
>
扁形动物门
> 吸虫纲
吸虫纲Trematoda都是营寄生生活的扁形动物,原始的种类主要寄生在软体动物、变温脊椎动物等的体表或腔隙。也就是主要为外寄生种类,例如单殖吸虫类(Monogenea)、盾腹吸虫类(Aspidogastraea)。进化的种类主要寄生在脊椎动物体内,为内寄生种类,例如复殖吸虫类(Digenea)。由于寄生生活方式,吸虫类在形态及生理上产生了相应的适应性特征;例如体表的纤毛及腺细胞退化或消失,出现了附着寄主组织的吸钩及吸盘;消化系统趋于退化,出现了兼有保护及吸收营养物质的皮膜;内寄生种类行无氧呼吸;神经感官的不发达;强大的繁殖能力及复杂的生活史等都与其寄生生活相关。但吸虫类的幼虫体表仍具有纤毛、并营自由生活,这一点与寄生的绦虫类不同。
吸虫纲
Trematoda
吸虫纲的主要特点
身体呈杆状,体表无纤毛,具腺细胞和
出现了外皮层(原生质的合胞体),可抵抗寄主体内消化酶的作用、吸收营养、进行气体交换
消化系统趋于退化,神经系统不发达,感官消失,仅某些幼虫阶段可出现眼点,适应短暂自由生活
具附着器,如口、腹吸盘、小钩或小刺
生殖系统发达,生活史复杂,有更换寄主的现象
单殖亚纲 Monogenea
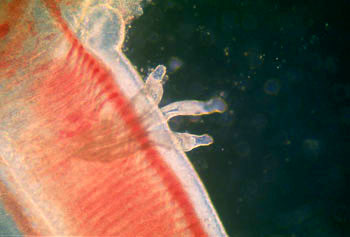
现在有些学者将单殖亚纲上升为纲与吸虫纲并列。本亚纲为体外寄生吸虫。生活史简单,直接发育,不更换寄主。主要寄生于鱼类、两栖类、爬行类等的体表和排泄器官或呼吸器官内,如鳃、皮肤、口腔,少数寄生在膀胱内。常缺少口吸盘,体后有发达的附着器官,其上有锚和小钩。眼点有或无。排泄孔一对,开口在体前端。仅有一个寄主,具钩毛蚴虫。
三代虫
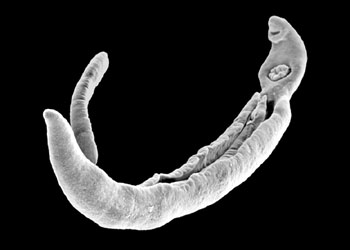
(Gyrodactylus)是侵害淡水鱼类(鲤、鲫、鳟等)的寄生虫,也寄生于两栖类。有20余种,淡水鱼场中常发现,使鱼类患三代虫病。三代虫身体扁平纵长,前端有两个突起的头器,能够主动伸缩,又有单细胞腺的头腺一对,开口于头器的前端。此虫没有眼点,口位于头器下方中央,下通咽、食道和两条盲管状的肠在体之两侧。体后端的固着器为一大形的固着盘,盘中央有2个大锚,大锚之间由2条横棒相连,盘的边缘有16个小钩有秩序地排列着。三代虫用后固着器上的大锚和小钩固着在寄主的身上,同时前端的头腺也分泌粘液,用以粘着在寄主体上或像尺蠖一样的慢慢爬行。 三代虫是雌雄同体,有卵巢2个及精巢一个,位于身体后部。三代虫为卵胎生,在卵巢的前方有未分裂的受精卵及发育的胚胎,在大胚胎内又有小胚胎,因此称为三代虫。 三代虫寄生于鱼类体表及鳃上,对鱼苗及春花鱼种危害很大,三代虫用锚钩钩住寄主的表皮组织,能造成体表的创伤,病鱼皮肤上形成灰蓝色无光泽的粘液膜。感染三代虫的鱼种初呈极度不安,狂游于水中,继则食欲不振、消瘦、以致死亡。三代虫繁殖最适温度为20℃左右,越冬鱼池内放养当年鲤、鲫都易感染;2年以上的鱼不会害病,但为本虫的带虫者。三代虫感染的方式主要靠接触传染,脱离母体的幼虫能在水中自由游泳寻觅寄主。
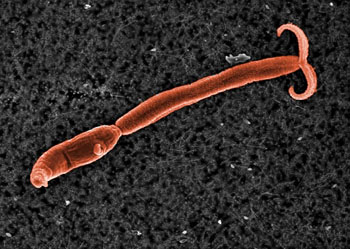
指环虫
(Dactylogyrus)虫体通常为长圆形,动作像尺蠖,寄生在各种鱼类的鳃上。身体前端有4个瓣状的头器常常伸缩,头部背面有4个眼点,在体后端腹面有一个圆形的固着盘,盘的中央有2个大锚,盘的边缘有14个小钩,在两大钩之间有1~2条横棒相连。口通常呈管状,可以伸缩,位于身体前端腹面靠近眼点附近,口下接一圆形的咽,咽下为食管,接着是分2支的肠,2条肠的末端通常在后固着盘的前面相连,使整个肠成环状,但也有不相连而呈盲管状的。 指环虫也为雌雄同体,有一个精巢和一个卵巢,卵大而量少,通常在子宫中只有一个卵,但能继续不断地产卵,所以繁殖率也相当高。卵产出后就沉入水底,经数日后即孵出幼虫,幼虫在水中游泳,遇到适当的寄主时附于其鳃上,脱去纤毛发育为成虫。 指环虫病通常发生在夏季,流行迅速。由于指环虫钩于寄主的鳃丝上,破坏鳃丝的表皮细胞,刺激鳃细胞分泌过多的粘液,使病鱼呼吸困难;由于鳃瓣表面受到破坏,附生各种细菌,严重感染时可引起鱼的死亡,特别是对草鱼的鱼苗和鱼种的危害性更为严重。预防三代虫指环虫病可在冬季进行清池、杀灭病鱼、引水入池后应静置几天,待病原生物死亡后,再放鱼苗鱼种,在放鱼种入池前可用高锰酸钾消毒,发现流行病时也可用此法洗浴治疗。
合体吸虫
(Diplozoonparadoxum)也是淡水鱼鳃部的寄生虫。是由两个虫体永远呈抱合状,故名合体虫。幼虫期一个个体的腹吸器吸着另一个个体的腹部突起,使两个虫体形成X形抱合。似多盘吸虫(Polystomoides)后吸器发达,其上有6个小的凹陷呈吸盘状,具一对钩。
三代虫(Gyrodactylus)为卵胎生,在卵巢的前方有未分裂的受精卵及发育的胚胎,在大胚胎内又有小胚胎,故名
三代虫(Gyrodactylus)寄生于淡水鱼类体表及鳃上,用锚钩钩住寄主的表皮组织,能造成体表的创伤,最终死亡
指环虫(Dactylogyrus)虫体通常为长圆形,动作像尺蠖,寄生在各种鱼类的鳃上,使病鱼呼吸困难甚至死亡
盾腹亚纲 Aspidogastraea
是吸虫纲中很小的一类,主要寄生在软体动物体表,与寄主的寄生关系很疏松,少数种寄生在鱼、海龟等脊椎动物的体表、消化系统及排泄系统。例如盾腹虫(Aspidogaster)身体腹面具有一个极大的腹吸盘,几乎占整个身体的腹面,吸盘上有纵行及横行肌肉将吸盘分成成行成排的许多小室(alveoli)。具口、咽、囊状肠。一个排泄孔开口在身体后端中央,生殖系统相似于复殖吸虫,大多数种类直接发育,少数种有幼虫期。寄生于脊椎动物的种类要求有一中间寄主。许多种类没有寄主的专一性,在软体动物及鱼体上均可生活及产卵。这一类似乎说明由自由生活到寄生生活的过渡类型。
复殖亚纲 Digenea
是吸虫纲中最大的一类,已报导的种超过了6000种,其中许多是人体及家畜等体内寄生虫,有很大的经济重要性。成虫有吸盘1个或2个,体后部无复杂的固着器,成虫无眼点,而幼虫有退化的感光器。生活史复杂,需要2个以上的寄主,终寄主为脊椎动物。其中间寄主为软体动物螺类、甲壳类、鱼类及水生植物等,具几个幼虫期。一般称为肠吸虫,例如布氏姜片虫;寄生在肝脏、胆管内的称为肝吸虫如肝片吸虫;寄生在血液中的则称为血吸虫。
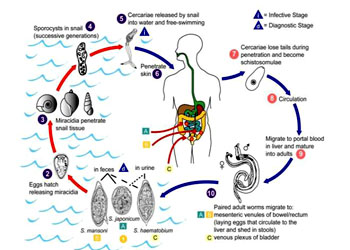
日本血吸虫(Schistosoma japonicum)成虫雌雄异体,雄虫体长约10~20mm,宽约0.5mm,
口吸盘
及
腹吸盘
均位于身体前端,腹面体壁内褶形成一沟称
抱雌沟
,雌虫即位于此沟内呈
雌雄抱合
状,寄生于人体或哺乳动物门静脉及肠系膜静脉内
雌雄成虫寄生在人肠系膜的小静脉中,在抱合中进行交配,在小静脉或肠壁的小血管里产
卵
,产出的卵部分随血流到达肝脏,部分卵沉积在肠壁,经十多天,卵内已发育成
毛蚴
(miracidium),它分泌溶组织酶,穿破肠壁落入肠腔,随寄主粪便排出体外
虫卵随粪便排出体外,一般存活时间不超过20天。从卵内孵出的
毛蚴
呈梨形,半透明,灰白色,周身被有纤毛,在水中近水面处作直线游动,有一定的趋光性和向上性。毛蚴的抵抗力较弱,在水中约存活1~3天。如遇到中间寄主钉螺则进入螺体内继续发育
钉螺
(Oncomelania hupensis)是日本血吸虫唯一的中间宿主,隶属于腹足纲盖螺科钉螺属。栖息于淡水水域但水陆两栖。个体小,尖圆锥形,壳高约10毫米,宽3~4毫米。壳面光滑或有粗、细纵肋。壳口呈卵圆形,外唇背侧有隆起唇嵴或无。肉可食。
当毛蚴遇到钉螺,即自钉螺软体部分侵入螺体,在螺体内先形成
母胞蚴
(mother sporocyst),经无性生殖产生许多
子胞蚴
(daughter sporocyst),子胞蚴再经无性生殖形成大量的尾蚴(cercaria),每头螺体内可多至数万条尾蚴
尾蚴
(cercaria)分体部和尾部,尾部是分类的重要特征,体部有吸盘及头腺。在有水的条件下,成熟尾蚴从钉螺体内逸出,一般密集在水面上,当接触人、畜皮肤(或粘膜)时,借其头腺分泌物的溶解作用及机械伸缩作用侵入皮肤,脱去尾部成为童虫
童虫
(schistosomula)先侵入小静脉或淋巴管,随血液循环经右心而到肺,经肺部毛细血管进入肺静脉入左心,进而经大循环流至全身各部,但只有到达肠系膜静脉的童虫才能发育成熟,也可以到肝门静脉内发育到一定程度后,最后仍回肠系膜静脉寄生
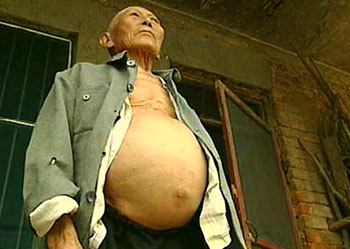
血吸虫病是热带与亚热带地区重要的传染病之一,晚期病人肝脾肿大硬化,以致出现腹水,腹部突出,俗称“
大肚子病
”,病人极端消瘦虚弱,晚期儿童患者可造成生长发育受到障碍,出现侏儒症、智力消退。晚期成年患者会出现生殖机能减退、严重病人最后死亡
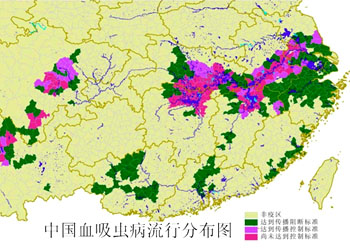
血吸虫病是严重危害我国人民健康的一种寄生虫病,其流行区分布于长江流域及长江以南广大地区。1972年在湖南马王堆出土的西汉古尸的肝脏中查见了日本血吸虫卵。防治血吸虫病需以防为主,灭螺、粪管、水管及预防感染。目前我国血吸虫病已基本得到控制
日本血吸虫
(Schistosoma japonica):是寄生在人体及哺乳动物静脉血管中的一种吸虫。寄生于人体的血吸虫共有3种,在我国流行的只有一种,即日本血吸虫(Schistosoma japonicum),是我国人体最重要的寄生虫之一。在我国主要分布于长江流域以南,包括台湾省共13个省市、347个县,有数亿人口受到威胁,另外日本、印度尼西亚、菲律宾等东南亚地区也是日本血吸虫病的重要流行区。另外两种是埃及血吸虫(S.haematobium),分布于非洲,亚洲西部,欧洲南部。曼氏血吸虫(S.mansoni),主要分布于非洲及拉丁美洲。日本血吸虫成虫雌雄异体,雄虫体长约10~20mm,宽约0.5mm,口吸盘及腹吸盘均位于身体前端,腹面体壁内褶形成一沟称抱雌沟,雌虫即位于此沟内呈雌雄抱合状。精巢(睾丸)7个排成一行,位于身体前端,肠分两支,后端汇合。雌性成虫较雄虫细长,体长12~26mm,宽约0.3mm,卵巢长圆形位于身体中部,卵黄腺位于卵巢之后,卵膜腔及子宫位于卵巢之前。子宫中常充满卵粒,每头雌虫每日可产卵1000~3000粒之间。雌雄成虫寄生在人肠系膜的小静脉中,在抱合中进行交配,在小静脉或肠壁的小血管里产卵,产出的卵部分随血流到达肝脏,部分卵沉积在肠壁,经十多天的发育,卵内已发育成毛蚴,它分泌溶组织酶,穿破肠壁落入肠腔,随寄主粪便排出体外。在水中毛蚴自卵壳出来,自由游泳。如遇到中间寄主钉螺(Oncomelania hupensis)则进入螺体内继续发育。在螺体内先形成母胞蚴,经无性生殖产生许多子胞蚴,子胞蚴再经无性生殖形成大量的尾蚴,在螺体内完成二代胞蚴,其发育时间在夏季约需2个月左右,冬季则时间更长,每头螺体内可多至数万条尾蚴。尾蚴成熟后由螺体内逸出,在水中自由生活,多浮于水面,1~2天内具有感染力。尾蚴甚至在潮湿的土壤,植物上的露水中亦能逸出,人如与含有尾蚴的水接触,则尾蚴靠头部的穿刺腺分泌溶组织酶及机械性作用,从皮肤或粘膜处侵入人体,侵入时尾部脱落,其尾蚴尾部分叉,是分类的重要特征。尾蚴进入人体后称为童虫,童虫先侵入小静脉或淋巴管,随血液循环经右心而到肺,经肺部毛细血管进入肺静脉入左心,进而经大循环流至全身各部,但只有到达肠系膜静脉的童虫才能发育成熟,也可以到肝门静脉内发育到一定程度后,最后仍回肠系膜静脉寄生。人体感染25天后雌虫开始产卵,35天后患者粪便中出现虫卵。虫卵也部分沉积在肝脏及结肠等处。成虫寿命数年,有的可达10~20年,个别病例最长的有30~40年的记载。人体受尾蚴感染后,几天内会出现皮肤的荨麻疹,也可出现速发型或迟发型皮炎。几周后由于大量童虫在体内移行会引起患者食欲不振、阵咳、呼吸受阻,甚至胸腹疼痛。如大量感染、2~3个月后患者发热、消瘦、粪便中有浓血;6~7个月后肝脾开始肿大、发热及便血症状消失,转入慢性期,这时大量的虫卵在肝脏及肠壁沉积形成囊肿,随后组织纤维化、肝脏开始硬化以致肿大。晚期病人肝脾进一步地肿大硬化,以致出现腹水,腹部突出,并伴随黄疸症状,病人极端消瘦虚弱,晚期儿童患者可造成生长发育受到障碍,出现侏儒症、智力消退。晚期成年患者会出现生殖机能减退、严重病人最后死亡。日本血吸虫病在我国长江流域及其以南地区的流行是受多方面的因素所影响,江南一带土壤肥沃、水草丰富、湖泊滩洲众多,为钉螺的繁殖提供了良好的孳生地。钉螺长约1cm,壳塔形,6~8个螺层,有厣、壳口外缘有一条凸起唇嵴,水陆两栖生活,钉螺的地理分布决定了血吸虫病的流行区域。另外目前的生产方式及生活方式也造成了它的流行,我国南方种植水稻、农田沟渠纵横,农民多用未加处理的生粪施肥。人们的生活用水也多来自河流、湖泊等未加处理的自然水。这样造成了钉螺的扩散,有利于虫卵的传播与孵化,也有利于毛蚴及尾蚴寻找寄主。各种家畜如牛、羊、猪、狗及一些野生脊椎动物也是日本血吸虫病的保存宿主。以上这些因素导致了日本血吸虫病的流行。1955年以后,在党与政府的重视之下,成立了“防治血吸虫病领导小组”及“全国血吸虫病研究委员会”领导血吸虫病的防治与研究工作。由于科技人员及广大群众的共同努力,经过积极地治疗病人及病畜、全面消灭钉螺、加强粪便管理、个人防护等多种措施,到1981年已有70%的流行县市基本上消灭了血吸虫病。但就全部流行区内消灭血吸虫病尚待今后的努力。
布氏姜片虫(Fasciolopsis buski)成虫是人体寄生吸虫中
最大
的一种,虫体平均长为30mm,宽为12mm左右,身体扁平,卵圆形,皮层有体棘,生活时肉红色,固定后为灰白色,体形像姜片,故名姜片虫
布氏姜片虫成虫在人或猪的肠腔中异体或同体授精,每头雌虫日产卵15000~25000粒,卵随寄主粪便排出体外并发育成毛蚴,遇中间寄主
扁卷螺
等即进入其体内发育成胞蚴,在螺体内发育一代胞蚴及二代雷蚴,最后形成尾蚴
尾蚴在水中从螺体逸出,遇适合的媒介植物如红菱,荸荠及茭白等水生寄主则附着其上,脱去尾部形成后尾蚴(囊蚴)。当人生食菱角及荸荠时即进入人体 、发育成成虫。病人感染姜片虫后,轻型者仅腹痛、消化不良、四肢及面部浮肿、贫血乏力,影响儿童发育
布氏姜片虫
(Fasciolopsis buski):成虫寄生在人及猪的小肠内。主要分布在亚洲温带及亚热带地区,我国多发生在长江以南、呈点状分布。成虫长圆形、肥厚多肉、长20~70mm、宽10~20mm,是寄生于人体的最大的一种吸虫。口吸盘及腹吸盘均位于身体的前半部,雌雄生殖孔分别开口在腹吸盘之前。肠分两支,在体侧呈波浪状弯曲。精巢一对,呈多分枝状,前后排列。卵巢一个呈三分枝状,位于身体中部精巢之前。卵黄腺位于体侧。成虫在人或猪的肠腔中异体或同体授精,每头雌虫日产卵15000~25000粒,卵随寄主粪便排出体外,在水中经3~7周发育成毛蚴,毛蚴存活1~2天,遇中间寄主扁卷螺即进入其体内发育成胞蚴。姜片虫的中间寄主螺有十多种,在我国主要的中间寄主是尖口扁卷螺(Hlppeutls cantorl),半球多脉扁卷螺(Polypylis hemisphaerula)及凸旋螺(Gyraulus convexiusculus)。在螺体内发育一代胞蚴及二代雷蚴,最后形成尾蚴,尾蚴在水中从螺体逸出,遇适合的媒介植物如红菱,荸荠及茭白等水生寄主则附着其上,脱去尾部形成后尾蚴(囊蚴)。当人生食菱角及荸荠时即进入人体 、发育成成虫。病人感染姜片虫后,轻型者仅腹痛、消化不良等症状。中度患者,腹痛腹泻、恶心呕吐、失眠症等。重度时四肢及面部均浮肿、贫血乏力,特别对儿童患者还会造 成生长发育的障碍。防治原则与日本血吸虫相似。
卫氏并殖吸虫(Paragonimus westermani):又称肺吸虫,成虫寄生在人及虎、狼、狗等动物的肺部。
淡水螺是肺吸虫的第一中间寄主,多属黑螺科(Melaniidae),如孵囊螺、短钩螺、黑螺(图)等
淡水虾或蟹类是肺吸虫的第二中间寄主,例如在我国南方多为溪蟹(图)、华溪蟹等,在我国东北地区主要是|蛄虾类
卫氏并殖吸虫
(Paragonimus westermani):又称肺吸虫,成虫寄生在人及虎、狼、狗等动物的肺部。我国主要分布在南方及东北地区。全世界已报道的肺吸虫有39种之多,我国有10多种,但致病性的主要是卫氏并殖吸虫。成虫卵圆形肥厚,长8~16mm、宽4~8mm、厚3~5mm。生活时呈红棕色。体表披棘,腹吸盘靠近身体中部、肠道两支也呈波浪状弯曲,精巢两个、亦分枝,但左右并列于身体后端,卵巢分瓣、与子宫也并列于身体中部,故名并殖吸虫。雌雄以共同的生殖孔开口在腹吸盘偏后侧。肺吸虫寄生在寄主肺内,在肺内形成囊肿,感染2~3个月后,童虫发育成熟。成虫寿命5~6年,每头雌虫日产量为10000~20000粒,产卵后囊肿破裂,卵随咳嗽及痰排出体外。卵在水中发育成毛蚴,遇第一中间寄主淡水螺则进入其体内,可被肺吸虫感染的淡水螺有许多种,多属黑螺科(Melaniidae),如孵囊螺(Brotia)、短钩螺(Semisulcospira)、黑螺(Melanoides)等。在螺体内经过一代胞蚴、二代雷蚴,发育成尾蚴。从感染螺体到尾蚴逸出约需2~3个月左右。尾蚴逸出后在水中自由生活,很快找到第二中间寄主淡水虾或蟹类,例如在我国南方多为溪蟹(Potamon)、华溪蟹(Sinopotamon)等,在我国东北地区主要是|蛄虾类(Astacidae)。尾蚴脱去球形尾部进入第二中间寄主后形成后尾蚴,后尾蚴是其感染阶段,人若生食感染尾蚴的虾蟹后,后尾蚴则脱去囊壁穿过寄主肠壁进入腹腔,再穿过横膈膜进入胸腔,在肺内寄生。童虫及成虫均有移行特性,并随时可停下来在寄主皮下、肝、肌肉、胸腔等处异位寄生。肺吸虫在人体引起的症状主要是咳嗽、痰中带血,由于后尾蚴、童虫及成虫有由囊中钻出、并在组织中移行的特性,所以症状常表现随寄生部位而不同,例如肠壁感染出现腹疼腹泻,如侵入脑,出现癫痫,寄生在淋巴组织可引起淋巴节肿大,寄生于皮下引起皮下结节等。
中华枝睾吸虫(Clonorchis sinensis)生活时呈肉红色,固定后灰白色,体内器官隐约可见,在虫体后1/3处有2个前后排列的树枝状睾丸,为该虫主要特征之一,故称枝睾吸虫
华枝睾吸虫的第一中间寄主为淡水螺,如
沼螺
;第二中间寄主主要是鲤科鱼类,如草鱼、青鱼、鳊、鲤、鲫、土鲮、麦穗鱼及米虾、沼虾等
中华枝睾吸虫(Clonorchis sinensis)成虫寄生在人、狗、猫等的
肝脏及胆管
内,患者感染后出现慢性腹泻、胆囊炎、黄疸、水肿、肝疼等,病情严重时常并发肝硬化、肝癌造成死亡
中华枝睾吸虫
(Clonorchis sinensis):成虫寄生在人、狗、猫等的肝脏及胆管内,过去曾在广东、福建一带及台湾地区流行,现南方诸省虽仍有报导,但多呈点状分布。成虫扁平、细长如叶状,长10~25m,宽3~5mm。腹吸盘较小在虫体前1/3处,肠为两支,由前向后直到身体后端,精巢(睾丸)两个、前后排列在虫体后2/3处。卵巢一个、细小分叶、位于身体中部与后1/3交界处。子宫管状细长,由卵膜腔开始盘旋而上,开口于腹吸盘前方。卵随寄主粪便排出体外发育为毛蚴,第一中间寄主为淡水螺。在我国有3个科7个种均可作为中间寄主,例如纹沼螺(Parafossarulus striatulus)、赤豆螺(Bithynia fuchsianus)等。在螺体内约需3个月完成胞蚴、雷蚴及尾蚴的发育。第二中间寄主为鲤科鱼类,如草鱼(Ctenopharyngodon idellus),青鱼(Mylopharyngodon aethiops)等,在其中形成后尾蚴,人取食未经煮熟的鱼而被感染。患者感染中华枝睾吸虫之后出现慢性腹泻、胆囊炎、黄疸、水肿、肝疼等症状,病情严重时常并发肝硬化、肝癌造成死亡。
肝片吸虫(Fasciola hepatica)又称羊肝蛭。成虫寄生在牛、羊及其他草食动物和人的肝脏胆管内,有时在猪和牛的肺内也可找到
肝片吸虫的中间寄主为
锥实螺
,毛蚴进入螺的肝脏后,进而发育成胞蚴、雷蚴,并无性生殖出尾蚴,在水中营自由生活
牲畜饮水或吃草时吞进囊蚴即可感染肝片吸虫。牛羊的肝脏胆管中如被肝片吸虫寄生,可引起肝炎、影响消化、贫血、消瘦及浮肿
肝片吸虫
(Fasciola hepatica):又称羊肝蛭。分布于世界各地,尤以中南美、欧洲、非洲等地比较常见。我国动物虽有感染,但人体感染少见。虫大,虫体长为20mm~40mm,宽5mm~13mm。体表有细棘,前端突出,略似圆锥,叫头锥。口吸盘在虫体的前端,在头锥之后腹面具腹吸盘。生殖孔在腹吸盘的前面。口吸盘的底部为口,口经咽通向食道和肠,在二肠干的外侧分出很多的侧枝,精巢2个,前后排列呈树枝状分支,卵巢一个呈鹿角状分支,在前精巢的右上方;劳氏管细小,无受精囊。虫卵椭圆形,淡黄褐色,卵的一端有小盖,卵内充满卵黄细胞。成虫寄生在牛、羊及其他草食动物和人的肝脏胆管内,有时在猪和牛的肺内也可找到。在胆管内成虫排出的虫卵随胆汁排在肠道内,再和寄主的粪便一起排出体外,落入水中。在适宜的温度下经过2~3周发育成毛蚴。毛蚴从卵内出来体被纤毛在水中自由游动。当遇到中间寄主锥实螺,即迅速地穿进其体内进入肝脏。毛蚴脱去纤毛变成囊状的胞蚴,胞蚴的胚细胞发育为雷蚴。雷蚴长圆形,有口、咽和肠。雷蚴破胞蚴皮膜出来,仍在螺体内继续发育,每个雷蚴再产生子雷蚴,然后形成尾蚴。尾蚴有口吸盘和腹吸盘和长的尾巴。尾蚴成熟后即离开锥实螺在水中游泳若干时间,尾部脱落成为囊蚴,固着在水草上和其他物体上,或者在水中保持游离状态。牲畜饮水或吃草时吞进囊蚴即可感染。囊蚴在肠内破壳而出,穿过肠壁经体腔而达肝脏。牛羊的肝脏胆管中如被肝片吸虫寄生,肝组织被破坏,引起肝炎及胆管变硬,同时虫体在胆管内生长发育并产卵,造成胆管的堵塞,影响消化和食欲;同时,由于虫体分泌的毒素渗入血液中,溶解红血细胞,使家畜发生贫血、消瘦及浮肿等中毒现象。人体感染可能是食生水、生蔬菜而得,因此在牧场中应改良排水渠道,消灭中间寄主锥实螺,禁止饮食生水、生菜,可使人免受感染。
吸虫纲的形态与生理
外形
:吸虫类的身体一般为或长或短的卵圆形、背腹均扁平、前端稍尖、后端稍钝。体长由1mm~7cm之间,身体无色或灰色或由于生殖腺及肠道中的食物而使身体呈现黄色、褐色或红色等。吸虫类显著的体表特征是具有附着器官,在不同的种类其附着器的结构是不同的。原始的单殖吸虫亚纲,主要是用发达的粘着器官附着,这种粘着器多由腺体构成,或由腺体及肌肉构成的盘,或是体壁凹陷或外突,其上有粘腺,而且附着器多出现在身体的一端或两端。如为两端则分别被称为前吸器(prohaptor)和后吸器(opisthaptor)。一些种在后吸器的中央还形成2~4个角质化的钩(hooks)或锚(anchors),边缘还有许多小钩,它们用粘液或吸钩附着在寄主体表,例如三代虫(Gyrodactylus)(图6-18A),还有的种粘着盘上出现一些凹陷,形成小的吸附器官,例如似多盘吸虫(Polystomoides)。凹陷之间也有放射棘相分隔,总之单殖吸虫的吸器上总是伴有钩、刺及角质化的网以起支持作用。在复殖吸虫类,粘着器官不发达,即使有粘着器也没有钩或锚伴随。它们的附着器官是与体壁分离由肌肉环绕的碗状吸盘,是靠吸盘的真空原理吸着在寄主体内,而不是粘着或钩着。一般具有两个,前端的称口吸盘(oral sucker),环绕在口的周围;另一个称腹吸盘(ventral sucker),在身体的腹面,例如中华枝睾吸虫(Clonorchis sinensis)。如果两个吸盘均存在时也可称双盘吸虫(Distome),如果后吸盘次生性的消失则称为单盘吸虫(monostome)。吸虫的口通常位于身体的前端,生殖孔位于身体的腹中线上。
体壁
:吸虫的体壁曾一度吸引着许多研究者的兴趣。过去一直认为体壁的最表层是一层角质层(cuticle),由深层的间质细胞分泌形成,是非生活物质。自1963年L.T.Threadgold用电子显微镜对肝片吸虫(Fasciolahepatica)的研究,从而否定了用光学显微镜研究所得出的结论,证明它是生活物质,并与深层的细胞有原生质相联,以后他又用电子显微镜对许多吸虫及其不同发育阶段的幼虫进行研究,结果还证实了它们的体壁中有线粒体(mitochondria)存在,这就更确定了吸虫的体壁表层是具有代谢活性的生活物质。吸虫体壁最外表的这层原生质层,没有细胞核及细胞膜,而是一层合胞体(syncytium),其中包含有线粒体及内质网(endoplasmic reticulum)、结晶蛋白所形成的刺以及胞饮小泡等,故称这层原生质层为皮层(tegument)。皮层基部为基膜(ba-sement membrane),只是它的细胞体及细胞核均下沉到实质中,由这些细胞伸出细长的管道(trabeculae)穿过肌肉层而与表面的原生质层相联。皮层的这种特殊结构及形态,不仅可以对抗寄主体内消化酶的作用,而且对吸虫通过体表吸收寄主的营养物质及进行气体交换都有重要的作用。吸虫由于寄生生活,皮层没有纤毛、杆状体等结构。皮层之下为肌肉层,外层为环肌、内层为纵肌;有的种还有斜肌,皮层与肌肉层也构成皮肌囊。肌肉层之内即为实质,身体两侧的实质中也有背、腹肌穿行。肌细胞亦为平滑肌,吸盘及咽处的肌肉特别发达。
营养
:吸虫类还保留有消化道,消化系统的结构很简单,包括口、咽、食道及肠,没有肛门。口周围常有吸盘围绕。咽壁具有肌肉便于抽吸食物、食道很短、肠为单枝或双分枝,其末端为盲端,也称为肠盲囊(intestinal caecum)。吸虫类均生活在有机质丰富的环境中,特别是内寄生的种类。随着寄生部位的不同,可以取食寄主的上皮细胞、血液、粘液等各种组织及组织的排出物,甚至可取食由寄主直接而来的食物颗粒。寄生在人体肝门静脉中的血吸虫(Schistosoma),直接以寄主的红血细胞为食。用体外培养实验证明:雄性血吸虫每小时取食30000个红血细胞;而雌性成虫是雄虫取食量的10倍,特别是在产卵阶段,一头雌虫每日产卵1000粒,要消耗掉其自身体重的10%,所以必须要从食物中获取大量的蛋白质及氨某酸。血吸虫取食后先行胞外消化,再行胞内消化。其胃层的上皮细胞形状可由柱形到扁平形,胃层细胞的基部含有非特异性酯酶,在血吸虫取食过程中,酶逐渐由基部移到细胞端部,当酶达到相当数量之后,细胞顶端破裂,酶被释放到肠腔与食物混合进行胞外消化过程;部分的血红蛋白的分解是在细胞内完成。代谢产物及不能被吸收的铁离子经过实质细胞再由排泄器官排出体外。在血吸虫的消化道中也发现了蛋白水解酶,它可以分解球蛋白成为肽的片段。吸虫类不仅可用消化道取食营养,其体壁的皮层对营养物质的吸收也具有重要的作用。许多溶解于虫体周围环境中的小分子物质可以通过吸虫类的皮层原生质膜的扩散作用(diffusion)从环境中吸收;环境中一些较大的颗粒吸虫类可以通过皮层的主动吸收(吞噬或胞饮)而获得营养。由于皮层内含有来自于自身的内源性酶(intrinsic enzymes)或来源于寄主的外源性酶(extrinsicenzymes),它可以从周围环境中通过其体壁而吸收营养物质。
呼吸与排泄
:吸虫类与涡虫相似,没有专门的呼吸器官。外寄生的吸虫类以及内寄生种类的营自由生活的幼虫阶段都是行有氧呼吸(aerobicrespiration),即通过体表进行气体交换。而内寄生的种类,由于周围环境中,特别是寄主的消化道内很少有游离的氧,故它们都是行无氧呼吸(anaerobic respiration)。无氧呼吸就是利用贮存在体内的营养物质――糖元(glycogen)在无氧条件下进行发酵作用以产生能量的过程。这个过程也称为糖酵解作用(glycolysis)。在酵解过程中一分子的葡萄糖借助于酶的作用氧化成两分子的丙酮酸盐,同时释放出两个高能磷酸键,然后再被还原成二分子的乳酸盐。在有氧条件下,一分子的葡萄糖也被氧化成二分子的丙酮酸盐,但不被还原成乳酸盐,而是经过三羧酸循环将丙酮酸盐进一步氧化成二氧化碳和水,在整个过程中释放出38个高能磷酸键。对比这两种异化过程,无氧呼吸是一种不完全的异化过程,它所释放出的能量仅为有氧呼吸的1/19,其代谢的终产物是乳酸盐(lactate)、醋酸盐(acetate)及丙酸盐(propionate)等中间产物,这是一种低效能的呼吸;而有氧呼吸是葡萄糖的完全氧化,其终产物为二氧化碳及水,并同时释放出更多的能量,比起无氧呼吸有氧呼吸则是高效能的呼吸。某些内寄生的吸虫,在有氧情况下也可以进行部分的三羧酸循环,但不能完成全过程。也曾有文献报导在一些吸虫的呼吸过程中,有着起重要作用的细胞色素系统,但在无氧呼吸中的意义还是不清楚的。吸虫类的排泄器官也是原肾,具两条排泄管及大量的焰细胞,单殖吸虫的排泄管分别开口在身体的前端,其代谢产物主要是氨,少数种类也可以产生尿素或尿酸,这些代谢产物在单殖吸虫类也可以通过体表排出,甚至是排泄的主要形式。另外其体内还含有较高浓度的氨基酸,如甘氨酸及脯氨酸等。可能原肾在体内主要是维持渗透平衡作用。在复殖吸虫类的一对排泄管在身体后端中央联合形成一个Y形膀胱(bladder),以单个肾孔开口在身体后端中央。在其原肾管液体中曾发现有氨、尿素等,因此具有排泄作用。内寄生种类由于体内、外环境中离子的浓度相对稳定,所以它的原肾对渗透调节作用是很有限的。这在许多内寄生的吸虫中得到证明。
淋巴系统
:一些复殖吸虫,如同口科(Paramphistomidae)及环肠科(Cyclocoelidae)的一些种,它们的实质中有管系统,由身体两侧的2―8个独立的管所组成,这些管分出盲枝分布到肠、生殖器官及吸盘等处,被认为是一种淋巴系统。这些管是由扁平的间质细胞组成,管内有液体,其中也漂浮着游离的细胞,整个淋巴系统似乎担任着食物、排泄物及气体的分散功能。
神经与感官
:吸虫类的神经结构与涡虫纲相似,包括一对脑神经节,脑神经节之间有横的神经纤维相联。由脑神经节向前、后分别通出3对纵神经索,其中腹神经索更为发达。一般在吸盘及咽处有更多的神经分布。吸虫类由于寄生环境的稳定性,特别是内寄生的种类,其感官很不发达。感官多存在于生活史中自由生活的幼虫阶段或存在于一些外寄生的成虫。眼点结构也很原始,仅有色素杯,此外在体表散布有感觉毛或感觉乳突,一般在头区较为丰富。感觉器或是由一个神经细胞末端具一根纤毛组成,或是由几个神经末端联合形成。它们担任着触觉、化学感觉及触流感觉等功能,复殖吸虫的感觉毛并不外露,因有体壁的皮层膜所包围。
生殖
:吸虫类绝大多数种类为雌雄同体,极少数为雌雄异体(如血吸虫)。生殖结构较复杂。雄性系统包括精巢(睾丸)、输精管、贮精囊及生殖孔等(图6-19)。精巢的数目、形状及位置因种而异,常作为分类的依据。例如单殖吸虫亚纲精巢常多个,只有一个输精管;复殖吸虫亚纲精巢多为2个、输精管两个,后端联合形成贮精囊,贮精囊常包在阴茎囊(cirrus sac)之中,贮精囊的后端形成射精管(ejaculatory duct)及阴茎(cirrus),射精管的周围还有前列腺包围,阴茎的末端以雄性生殖孔开口在生殖腔中或单独开口在体外或由生殖腔中外伸出来。雌性生殖系统包括一个卵巢,由卵巢通出一条输卵管,输卵管后端接受受精囊管、卵黄腺管,三个管汇合后膨大形成卵膜腔(ootype),卵在卵膜腔中受精。卵膜腔周围围有梅氏腺(Mehli’sgland),离开卵膜腔后为一盘旋的管状子宫,子宫末端或开口在生殖腔或直接开口体外,生殖腔有肌肉包围,肌肉的收缩使卵易于排出。许多吸虫输卵管在进入卵膜腔之前,具有一短管,称为劳氏管(Laurer’s canal),其机能不清楚,可能是退化的阴道,为贮存过多的精子之用。吸虫在通常情况下行异体受精,有时也发生自体受精。交配时,雄性的前列腺分泌粘液,以保护精子的存活,交配后精子经雌性子宫上游,最后进入受精囊并贮存在其中。卵由卵巢排出后,经输卵管到卵膜腔或在进入卵膜腔之前与从受精囊出来的精子相遇而使卵受精。吸虫的卵也是外卵黄卵,受精卵周围接受卵黄物质,梅氏腺的功能是对卵壳的形成起模板作用或刺激卵黄细胞释放卵黄物质以及活化精子,也有人认为它的分泌物有滑润作用,以利于卵通过子宫。卵经过子宫、生殖孔之后排出体外。与寄生生活相关,吸虫具有巨大的产卵量,与自由生活的涡虫相比,有人估计吸虫的产卵数是涡虫的10000~100000倍之多。
生活史
:吸虫的生活史在不同的种类其复杂程度不同。单殖吸虫亚纲生活史简单,只有一个寄主,由成虫产卵后发育成钩毛蚴虫(onchomiracidium),钩毛蚴体表披有纤毛,在水中自由游泳生活一段时间。一般通过化学引诱寻找寄主――鱼,找到寄主后,在附着处如鱼的体表、鳃、鳃腔等部位脱去纤毛,然后变态成成虫。复殖吸虫生活史复杂,均有寄主更换现象。一般有2―4个寄主;两个自由生活的阶段,即寄主转移时期;生活史中有有性生殖及无性生殖时期。在2~4个寄主中只有一个寄主称终寄主(definitive host),吸虫在终寄主通常是脊椎动物体内行有性生殖。其他的寄主均称中间寄主(intermediatehost),吸虫在中间寄主体内均行无性生殖,通过无性生殖完成一个或几个世代。中间寄主通常是软体动物的螺类或其他无脊椎动物及植物。换句话说凡行有性生殖的寄主称终寄主,行无性生殖的寄主称中间寄主。不同种类的复殖吸虫发育过程中所经历的虫态期大致可分为卵(egg)、毛蚴(miracidium)、胞蚴(sporocyst)、雷蚴(redia)、尾蚴(cercaria)、后尾蚴(metacercaria)及成虫(adult)。但每种的吸虫所经历的幼虫期并不相同,每种幼虫无性繁殖的代数也有区别。
卵(egg)
吸虫卵大多呈卵圆形,多数具卵孔,有卵壳包围,由母体产出的卵,多数种在其卵壳内已发育成胚胎或小的毛蚴。卵的大小、形态及卵壳有种的特异性,常用来做鉴别及诊断成体类别的根据。卵的孵化受外界不同因素影响如水、温度、气压、酸碱度及光照都是影响孵化的因素。例如卵在10~37℃、pH4.2~9之外则停止发育,也有的种,卵是被寄主吞食之后才孵化,这时寄主体内的酶及CO的分压是其孵化的重要因素。
毛蚴(miracidium)
毛蚴是生活史中第一个自由游泳的时期,也是感染中间寄主的阶段。身体呈梨形、全身披有纤毛。例如日本血吸虫的毛蚴体长仅80~120μm。身体前端具有顶腺(apicalgland)和头腺(frontal gland),还具有神经节、眼点、化感器和触觉感受器等,身体两侧有一对原肾,中央有成团的生殖细胞。毛蚴很活跃,在水中自由游泳,以寻找中间寄主。有人认为寄主螺分泌的粘液对毛蚴有化学吸引力,使毛蚴易于找到寄主。也有人认为是由于毛蚴自身的运动使它寻找寄主,毛蚴的年龄决定了它寻找寄主的成功与否,因孵化后2~5小时的毛蚴生命力最强,运动范围大易于寻找寄主,超过一定年龄后毛蚴则失去活力,很难找到中间寄主。其他的理化因素,特别是光也有利于吸引它们寻找中间寄主。当毛蚴发现寄主螺之后,靠头部腺体分泌粘液帮助它穿透表皮组织进入螺体内,并同时脱去纤毛,进入胞蚴期。
胞蚴(sporocyst)
毛蚴进入螺体后,脱去纤毛成一囊状物,这就是胞蚴。除了毛蚴期的体壁、原肾保留之外,其他结构消失。胞蚴多寄生在螺体的足、鳃、触角、淋巴等处,靠体表吸收寄主的营养以供体内胚细胞的生长发育,一个胞蚴经过无性生殖方式可以产生许多雷蚴或子胞蚴。胞蚴期是吸虫行无性生殖阶段。
雷蚴(redia)
由胞蚴体内经繁殖出来的小个体即为雷蚴,体长圆形,后端较钝。具有口、咽及不分枝的肠。雷蚴很活跃,在螺体内爬行,常迁移到螺的肝区及生殖腺处,它们通过肠道或体表吸收寄主的营养。雷蚴也是吸虫类进行无性生殖、扩大种群数量的阶段。由雷蚴经过无性生殖可形成许多尾蚴或子雷蚴。一般雷蚴在螺体内繁殖多代,在螺体内达到相当数量之后才出现尾蚴。一些种类的雷蚴期也可进行寄主的转移。
尾蚴(cercaria)
尾蚴是吸虫生活史中第二个自由生活的阶段,绝大多数吸虫的尾蚴有一长形尾,有口、口吸盘,肌肉质咽及分枝的肠道。身体前端有穿透腺,其分泌物帮助尾蚴穿透寄主皮肤,也有原肾,其焰细胞的数目及排列是分类及鉴定种属的重要依据之一。尾蚴成熟后,自雷蚴体内出来,并离开中间寄主在水中自由游泳,寻找新的寄主(第二、三中间寄主或终寄主)。故尾蚴是吸虫的第二个寄主转移阶段。尾蚴对光有正趋性,游泳时多漂浮到水面,静止时沉入水中,它在水中的感染力也有一定的时间限制,超过最佳活力阶段,则很难进入新寄主体内。许多实验证明寄主体内的脂肪酸对尾蚴具吸引力。
后尾蚴(metacercaria)
尾蚴在水中接触到新寄主之后,脱去尾部靠头部的穿透腺进入寄主表皮内、分泌外壁形成胞囊状,称为后尾蚴(或囊蚴)。有的吸虫尾蚴进入第三中间寄主(啮齿类),在其体内发育成后尾蚴,例如重翼吸虫(Alaria)。
成虫(adult)
一旦尾蚴或后尾蚴进入终寄主之后或在寄主皮肤上脱去尾部(尾蚴),或在寄主消化道内脱去囊壁(后尾蚴),再经过体腔或肝门静脉移行到寄生部位,如肝、肺、及血液等处变态成成虫。不是所有的吸虫都经过上述各发育阶段,不同的种有不同的改变,最常见的改变是没有胞蚴或没有雷蚴;多于一代胞蚴或雷蚴;没有后尾蚴,由尾蚴直接感染终寄主。