水螅纲
钵水母纲
珊瑚纲
你的位置:
首页
>
动物界
>
刺胞动物门
> 钵水母纲
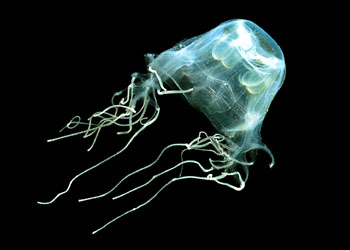
钵水母纲(Scyphozoa)生活史的主要阶段是单体、水母型,其水螅型阶段不发达、或完全消失。钵水母纲的水母体不同于水螅纲的水母体,这种区别主要表现在:钵水母纲的水母体一般体型较大,没有缘膜;胃循环腔复杂,辐射管发达,有内胚层起源的胃丝,胃丝上有刺细胞;中胶层中有外胚层起源的细胞及纤维;生殖细胞起源于内胚层,水螅纲水母均来源于外胚层;神经感官较发达,集中形成4~8个感觉器官。本纲已知约200余种,全部海洋生活,可分为5目。
钵水母纲
Scyphozoa
钵水母纲的主要特征
全部海产
世代交替,但水螅型退化或无,以水母型世代为主
单个,多为大型水母,中胶层厚,构造复杂
神经感官较发达,具有触手囊,具有眼点、平衡石、嗅窝等结构,有感光、平衡、化学感受器等功能
无缘膜,是钵水母与水螅水母的主要区别
消化循环腔复杂,辐射管发达,起源于内胚层的胃丝具刺细胞,所以钵水母内、外胚层都有刺细胞
生殖腺起源于内胚层
立方水母目 Cubomedusae
整个伞部呈立方形,伞缘也呈四边形,在间辐区由伞缘伸出一条或多条触手,触手在基部形成足叶,具有8个触手囊,位于间辐及正辐区伞缘上。下伞面向内延伸形成假缘膜(Velarium)。具有边缘神经环,并与触手囊相联,这与水螅水母有些相似。发育中钵口幼虫可以直接形成成体,主要分布在热带及亚热带浅海,少数在开阔的海洋中营漂浮生活。触手上的刺细胞具有很高的毒性,对浅海游泳者构成威胁,常见的代表种如手曳水母(Chiropsalmus),盒水母(Carybdea)等。
手曳水母(Chiropsalmus quadrumanus)
盒水母(Carybdea sivickisi)
罗氏箱水母(Carybdea robsoni)
冠水母目 Coronatae
水母体呈圆屋顶形、锥形、或扁平行,但外伞中部有一紧缩沟,将伞分成上、下两部分,相应的胃囊也被分成上、下两部分。沟下有一圈厚的足叶(pedalia),其下端是触手,伞缘具触手囊,主要为深海产,例如缘叶水母(Periphylla)。
缘叶水母(Periphylla periphylla)
缘叶水母(Periphylla periphylla)
缘叶水母(Periphylla periphylla)
十字水母目 Stauromedusae
是营固着生活的钵水母类,身体由水螅型及水母型联合形成,形如倒置的喇叭。其上伞面延长成柄状,末端具基盘,用以固着,下伞面向上,伞呈杯状,伞缘有8个边,有8簇短的触手,没有触手囊,仅在触手丛内各有一感觉小体,口四边形,位于下伞的中央,口周围有4个小的口叶,胃腔内有胃囊及隔板。浮浪幼虫没有纤毛,经爬行后,固着发育成成体。常分布在较冷的海水中,附着在海藻上生活,例如喇叭水母(Haliclystus)及高杯水母(Lucrenaria)等。
十字佐氏水母(Sasakiella cruciformis)
喇叭水母(Haliclystus borealis)
耳喇叭水母(Haliclystus auricula)
旗口水母目 Semaeostomae
是本纲中最常见的一个目。伞呈碗形、蝶形,伞缘有8个到许多个缺刻,缺刻中有触手囊。触手的数目、分布、形状随种而异。具口腕,口腕中有纤毛沟,由胃囊伸出复杂的辐射管,环管或有或无。典型的生活史是水母体→浮浪幼虫→钵口幼虫→横裂体→碟状幼虫→水母体。绝大多数在沿海生活。常见的种有海月水母(Aurelia)、游水母(Pelagia)、霞水母(Cyanea)等。海月水母(Aurelia aurita)营漂浮生活,体为白色透明的盘状,在伞的边缘生有触手,并有8个缺刻,每个缺刻中有一个感觉器,也称触手囊,囊内有钙质的平衡石(statolith),囊上面有眼点(ocellus),囊下面有缘瓣(lappet),缘瓣上有感觉细胞和纤毛,另外有2个嗅窝。当水母体不平衡时,触手囊对感觉纤毛的压力不同,而产生不平衡的感觉。在内伞的中央有一呈四角形的口,由口的四角上伸出 4条口腕(oral lobe)。消化循环系统比较复杂,由口进去为胃腔,位于体中央,向四方扩大成四个胃囊,由胃囊上和胃囊之间伸出分枝的和不分枝的辐管(radial canal),这些辐管均与伞边缘的环管(ring canal)相连。水流由口进去至胃腔,经过一定的辐管至环管,然后再由一定的辐管流至胃囊,经口流出。在胃囊的里面,有四个由内胚层产生的马蹄形的生殖腺,位于胃囊底部的边缘,在生殖腺内侧,长有很多丝状的结构称为胃丝,也是由内胚层形成的,其上有很多刺细胞,食物进入胃囊后,即被刺丝杀死,经消化后(细胞内和细胞外消化)由辐管分布到全身各部,胃丝也起着保护生殖腺的作用。 由生殖腺产生精子或卵(钵水母为雌雄异体),精子成熟后随水流至雌体内受精,也有的在海水中受精,受精卵经完全均等卵裂形成囊胚,再由内陷方式形成原肠胚,此时胚胎表面长出纤毛,成为浮浪幼虫,在海水中游动一个时期后,附于海藻或其他物体上,发育成小的螅状幼体(hydrula),有口和触手,可营独立生活,然后进行横裂,由顶而下分层为钵口幼体(scyphistoma),再进行连续横分裂形成一个个碟状个体,称横裂体(strobila)。横裂体成熟后一个个依次脱落下来,称为碟状幼体(ephyra),由它发育成水母成体。由此可见,钵水母的生活史虽有世代交替,但水母型发达而水螅型则退化成为幼虫。也有的钵水母无世代交替现象,只有水母型。 发状霞水母(Cyanea capillata)国内分布于黄海南部、东海北部;国外分布在日本。大型个体,伞直径为700~850毫米。伞部扁平呈圆盘状,外伞中央具许多刺胞。8个感觉器位于8个浅凹缘之间的底鄙,感觉棒梭形,其远端具一豆形平衡囊,没有眼点。下伞的纵辐位上,有8束排列U字形的触手,每束触手数日很多,通常较粗大的触手近于伞中,细小的靠近伞缘,下伞具辐肌束与环肌束。口腕发达,其长度超过伞半径。生活时体呈乳白色或带褐色。营浮游性生活。常群体出现在海面上,以幼小生物为食。繁殖期多在夏季。大量出现时会阻塞渔网,减少海水过滤,致使网衣破裂,影响渔业生产。另一些大型个体螫刺人体后,可造成严重创伤。
海月水母(Aurelia aurita)
夜光游水母(Pelagia noctiluca)
狮鬃水母(Chrysaora melanaster)
北极霞水母(Cyanea arctica)是最大的水母,伞径达2米
发状霞水母(Cyanea capillata)
发状霞水母(Cyanea capillata)
根口水母目 Rhizostomae
接近于旗口水母,但伞缘无触手,口腕愈合,口封闭,又形成了许多小的吸口。早期发育中具有正常的口,并有4个口叶,以后发育中4个口叶分枝成8个口腕,口腕再分枝愈合,原来口腕中的纤毛沟愈合成小管及吸口(suctorial mouth)。吸口、小管与胃腔相连,胃腔中也有辐射管,环管或有或无,具触手囊,代表种如海蜇(Rhopilema)、硝水母(Mastigias)等。海蜇含有丰富的维生素B,为名贵食品,我国东海、南海等沿海产量较丰富。
硝水母(Mastigias)爆发
巴布亚硝水母(Mastigias papua)
海蜇(Rhopilema esculenta)
钵水母的基本结构
:钵水母纲的水母体体型较大,伞缘直径一般在2~40cm之间,个别大的种直径可达1~2m,例如一种霞水母(Cyanea capillata)。伞呈圆盘形,如海月水母(Aurelia),或呈锥形、半圆形、蝶形等因种而异。由于体内的生殖腺或其他胃囊等结构具有色泽,而使身体在透明中出现局部的粉红色、桔红色等。身体也区分成上伞面及下伞面,无缘膜、伞缘具有一圈触手,不同的种触手的数目不同,触手或实心或空心,或长或短,也有少数种没有触手,例如根口水母类(Rhizostomae)。伞缘具有感觉器官,也称为触手囊(rhopalium),数目为4或4的倍数,海月水母有8个,呈缺刻状,因此将伞缘分成了8片。下伞中央的垂唇末端向外延伸,形成4个或8个口腕(oralarms),口腕向中心的一侧有沟。口腕具捕食的功能,取食时微小的食物可沿口腕沟进入海月水母(Aurelia)中,根口水母类口腕愈合,垂唇末端的口封闭,而形成许多新的细小的吸口,用以吸食。一般在触手、垂唇、口腕及伞的外表面分布有刺细胞。一些种类的下伞面生殖腺区向内凹陷形成生殖下窝(subgenitalpit),其功能不详,可能与动物的呼吸有关。钵水母类的中胶层很发达,也是由蛋白质及粘多糖形成的凝胶,其中含有胶原纤维。不同于水螅水母的是中胶层中游离着外胚层起源的变形细胞,这些变形细胞对动物的再生、组织修复起重要作用。中胶层也有很大的弹性,由于它能维持及调节离子的成分及浓度,而使身体在海水中保持一定的浮力。钵水母类的肌肉及运动相似于水螅水母,围绕着下伞缘有由外胚层形成的环行肌肉,触手上有纵行的肌纤维,它们的收缩造成水母的运动。
钵水母的消化
:钵水母类的胃循环腔比水螅水母复杂,原始的种类由口经垂唇进入中央的胃腔,胃腔向外延伸形成4个胃囊,胃囊之间有隔板(septum),隔板上有小孔,可使胃囊之间互相沟通以帮助液体的循环流动。隔板上有隔板肌,内缘有内胚层起源的胃丝(gastric filaments),共上含有许多刺细胞及腺细胞,可以固定及杀死进入胃腔的食物。例如十字水母类(Stauromedusae)就具有这种胃腔。较进化的钵水母类,例如海月水母,这种胃囊及隔板的结构仅在幼年阶段出现,成年阶段时形成了发达的胃腔及环流管系统。海月水母的胃环流管包括由口腕方向伸向伞缘的4条分枝的正辐管(perradial canals),由胃囊方向伸向伞缘的4条分枝的间辐管(interradialcanals),及位于正辐管与间辐管之间的8条不分枝的从辐管(adradial canals)。这些放射管在伞缘处均与环管相连。钵水母类均为肉食性动物,以小的甲壳类、浮游生物等为食,实际上也是一类悬浮取食者,它们以触手过滤水中的微小的浮游生物,经口腕沟靠纤毛作用送入口及胃腔,胃丝上的刺细胞杀死捕获物,再由胃丝上的腺细胞分泌消化酶消化食物,消化后的营养物靠环流管壁的纤毛摆动以推动营养物由胃腔经从辐管进入环管,再经正辐管、间辐管、胃腔及口将未消化吸收的食物残渣排出体外。
钵水母的神经与感官
:钵水母类的神经结构也是由外胚层形成的神经网,也具有突触传导。原始的种类例如立方水母类(Cubomedusae),也像水螅水母一样,在伞缘具有两个神经环。但多数的钵水母类已不存在这种伞缘神经环,而是神经细胞集中,形成4个或8个神经节分布在伞缘的触手囊中。显然,其中含有起博点神经元,因为如果切去全部的神经节,水母则失去博动的能力,如果切去部分神经节,甚至只留一个神经节,水母仍能做有节奏的收缩运动。所以神经节中含有起博点神经元,它控制着水母类的肌肉收缩运动。钵水母类伞缘的4个或8个触手囊也是它的神经感觉中心,它具有感光、重力感受及化学感受的功能。触手囊是由环管向外延伸形成一个中空的小盲管,其末端具有内胚层分泌的钙质颗粒,称平衡石(statolith)。外伞缘在平衡囊上端延伸形成笠(hood),用以保护及遮盖下面的平衡囊,其两侧有感觉瓣(sensory lappets),其上有感觉细胞及纤毛。当身体倾斜时,端部的结石与感觉纤毛受到刺激而引起运动以调节身体的平衡。另外,在触手囊上还有外胚层形成的小眼(ocellus),原始的小眼仅是表皮细胞内陷形成的一个小窝,其中分布有色素及感觉细胞,复杂的小眼有网膜状的感觉细胞及晶体,如立方水母类,对光线表现出正趋性。此外,在触手囊上、下伞面有一个表皮内陷形成的外,内感觉窝(sensory pit),是其化学感受器。钵水母类的触手囊具有敏锐的感觉能力,例如它能感受到比声波还微弱得多的次声波。有时风平浪静的海面会见到水母类的聚集或成群游动,有经验的渔民及海员会意识到几小时之后,海面将会有风暴的来临,这是由于空气中的气流及海浪的磨擦所产生的一种人不能查觉的次声波,而水母类能感受,并提前开始了迎战风暴的准备。因此人们把某些钵水母类看做是一种有效地预测风暴的指示生物了。仿生学家也利用了它的触手囊结构,成功的制成了风暴预测器,能提前十几个小时成功的预报风暴的来临、方向、级别等。为航海者提供了可贵的资料。
钵水母的生殖和发育
:钵水母类为雌雄异体,生殖腺位于胃襄内,由内胚层产生。原始的种类在隔板两侧共有8个生殖腺。无隔板的种类具有4个,海月水母就有4个马蹄形的生殖腺体在胃囊的底部,性成熟时常有色泽,易于识别。生殖细胞排到海水中或口腕处受精,受精卵经囊胚期发育成浮浪幼虫,经过一段自由游泳之后,用其前端固着在物体上发育成水螅型幼虫,称为钵口幼虫(scyphistoma),钵口幼虫以后由顶端到基部进行横裂生殖(strobilation),这时称为横裂体(strobila),以后横裂体由顶端开始依次脱离母体形成幼年的水母型体,称蝶状幼体(ephyra),横裂体可以生活一到数年,全部横裂体脱落之后,它又可以重新形成钵口幼虫,所以横裂体是无性生殖阶段。蝶状幼体很小,边缘有很深的缺刻。它经过大量的取食后发育成水母型成体。远洋漂浮的钵水母类没有固着生活的阶段,它们或是直接发育,例如游水母(Pelagia),棕色水母(Atolla),或是幼虫保持在亲本的胃腔内或其他部位,例如霞水母。