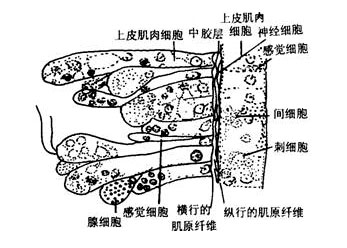
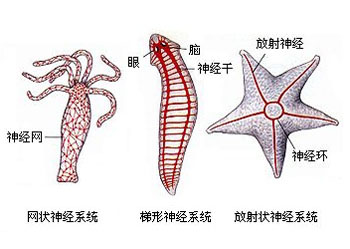
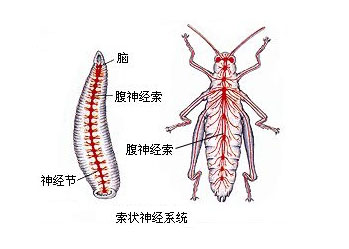
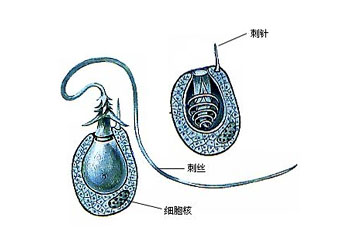
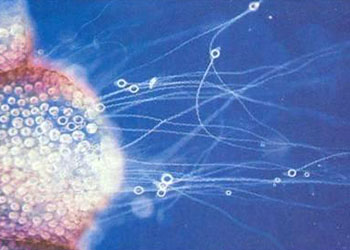
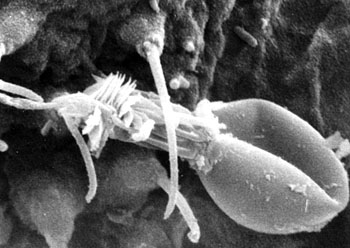
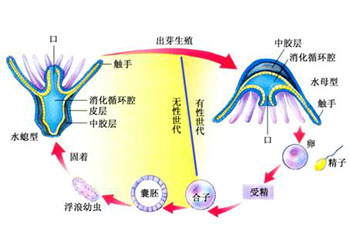
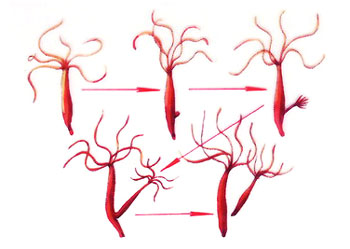
刺胞动物门
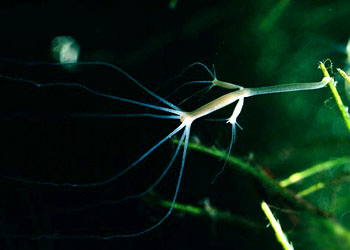
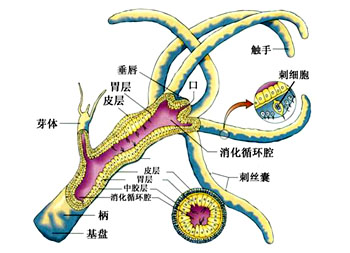
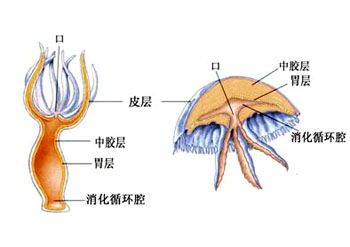
肌肉与运动:刺胞动物很少能做主动的移位运动,运动的能力是很有限的,特别是水螅型。运动是由表皮肌肉细胞中肌原纤维的收缩所引起。例如水螅的身体可做伸缩运动,伸展时体长可达15~20mm,收缩时体长仅0.5mm,这种伸缩是爆发式的,每5~10分钟左右爆发一次,主要是由外皮肌细胞的纵行肌原纤维的收缩所引起。身体一侧的肌原纤维的收缩可引起身体的弯曲。有时靠弯曲身体及触手并与基盘的交替附着而做翻斛斗式的运动。基盘处粘细胞可分泌大量的气泡,可使水螅在水面上做短暂的漂浮。钵水母类和珊瑚类的肌原纤维已与表皮细胞分离形成独立的一层肌纤维,例如水母的肌原纤维在下伞面及伞缘形成薄薄的一层肌肉环,有的被辐管分离成片状,其纤维有横纹,它们做有规律的收缩,使伞面有节奏的收缩运动。当伞缘收缩时,伞缘内的水被喷出,由于反作用力而推动水母体向上运动,当伞及伞缘肌肉舒张,被压缩的中胶层的弹性,使伞又恢复了原形,水又重新进入伞缘内,身体下沉,但由于收缩比舒张要快,所以水母还是可以向上垂直运动。水母类的垂直运动在有缘膜的水母可以看的更为清楚。一旦肌肉停止收缩,水母就会自然下沉。其水平方向的运动多是被动的,多是由于水流及风力所推动。又例如在珊瑚类,海葵的肌原纤维在隔膜上形成发达的牵缩肌,它的收缩非常有力,所以当海葵固着在岩石上是很难将它取下的。