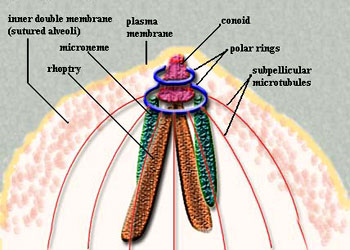
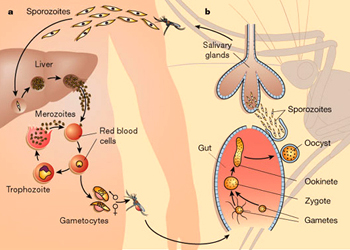
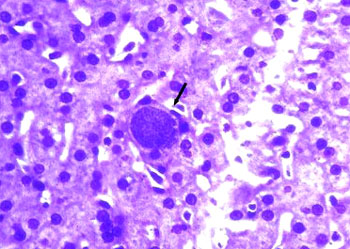
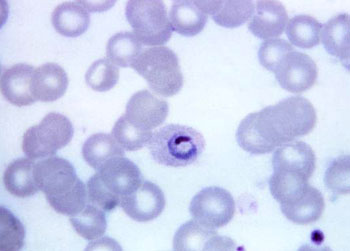
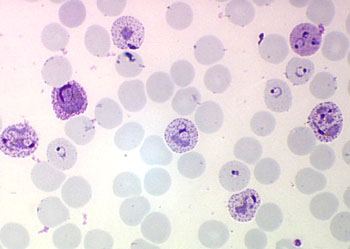
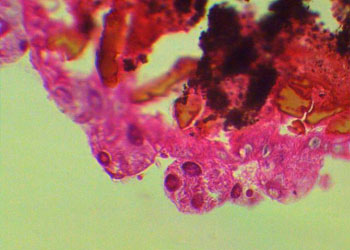
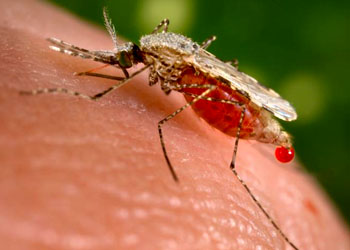
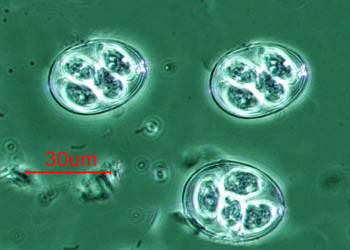
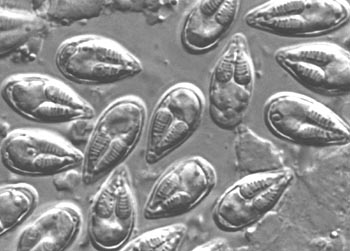
孢子纲Cestoda营寄生生活,没有运动细胞器,但在生活史中一定阶段,具有伪足或鞭毛。异养。生活史包括无性世代(裂殖生殖)和有性世代(孢子生殖)交替,寄主1~2个。很多具有顶复合器结构,有时将这类独立为顶复合门Apicomplexa,同时微孢子虫、囊孢子虫和粘孢子虫也分别独立为门。约有3500种,分布广,如疟原虫Plasmodium(疟疾,按蚊传播)、球虫Eimeria、焦虫Piroplasm、碘孢虫Myxobolus等。点击更多
孢子纲