植鞭亚纲动鞭亚纲
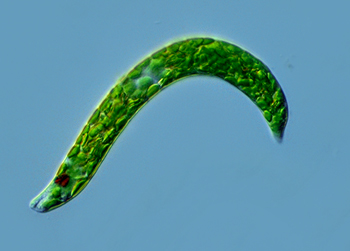
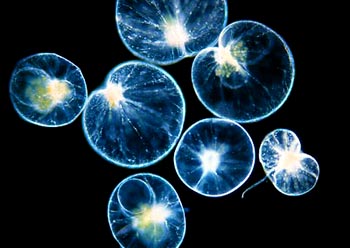
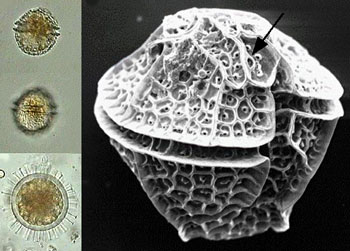
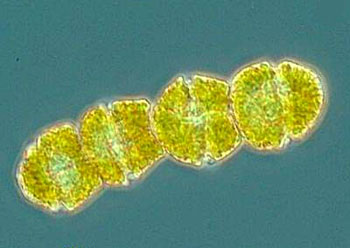
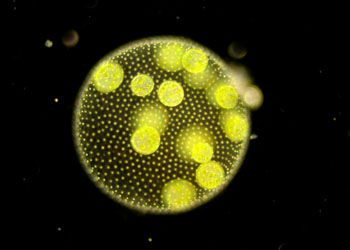
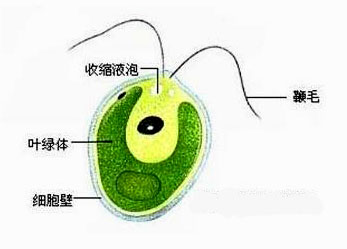
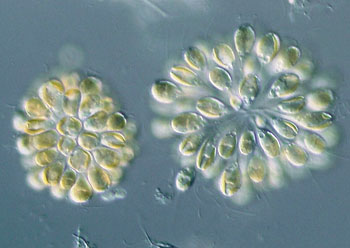
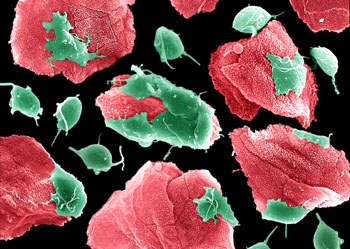
鞭毛纲(鞭毛虫纲)Mastigophora约10000种。一般具鞭毛,以鞭毛为运动器,鞭毛通常有1~4条或稍多,鞭毛内周缘部排列9条双联体微管、中央有2条中央微管。无性繁殖一般为纵二分裂,有性繁殖为配子结合或整个个体结合。在环境不良的条件下一般能形成包囊。根据营养方式的不同,可分为两个亚纲:植鞭亚纲Phytomastigina一般具有色素体能行光合作用,自由生活在淡水或海水中(自养),种类多,形状各异,如眼虫Euglena、盘藻Gonium、团藻Volvox、夜光虫Noctiluca(腰鞭毛虫类,可引发赤潮)、钟罩虫Dinobryon、尾窝虫Uroglena、合尾滴虫Synura。动鞭亚纲Zoomastigina无色素体,寄生、腐生或吞噬营养(异养),如利什曼原虫Leishmania(黑热病,白蛉子传播),锥虫Trypanosoma(昏睡病)、隐鞭虫Cryptobia(寄生于鱼鳃)、披发虫Ttrichonympha(共生于白蚁肠道,使其可以消化木材的纤维素)。点击更多
鞭毛纲