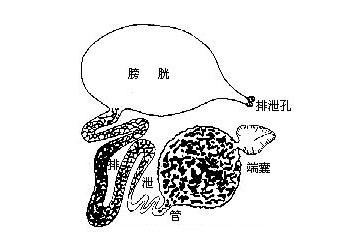
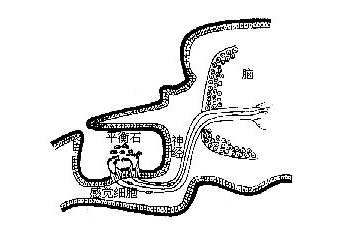
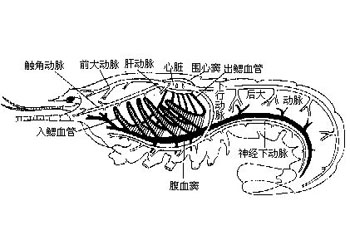
体壁结构与其他节肢动物相似,只是某些甲壳类如虾、蟹的外骨骼有相当程度的钙化,主要是因为上表皮及外表皮均有相当多钙盐的沉积。另外在表皮细胞的下面有皮下腺(tegumentalgland)和色素细胞(ehromatophores)。皮下腺是一群具分泌能力的腺细胞,并有长管穿过外骨骼开口在上表皮的表面,它的功能还不十分清楚。色素细胞是位于表皮细胞下面的结缔组织中的一些呈放射状分支的细胞,放射状的突起不能伸缩,但细胞中含有大量的色素颗粒,呈白、红、黄、蓝、褐、黑等色。根据生理状况的不同,颗粒或分散在整个细胞,或集中在细胞中心。其中红、黄、蓝色是类胡萝卜素,来自于食物,在生活状态时,这些色素常与蛋白质呈结合状态而表现出不同的颜色。当虾、蟹加热煮熟时,体表出现美丽的红色,这是因为加热使蛋白色沉淀出来而留下红色化合物――虾青素(astaxanthin)的缘故。事实上,一个单个的色素细胞可以包括一种到几种不同颜色的色素,任何一种色素都可独立移动。多色素的色素细胞仅出现在虾中。体色的改变有两种形式。一种是形态变色(morphologicalcolorchange),它涉及色素细胞中色素的丢失或形成,或是由于动物长期在一固定的环境及光照条件下细胞数目的改变而形成。另一种形式是生理变色(physiological color change)它是指对环境的迅速的颜色适应,来自于色素细胞中色素颗粒的分散与集中。最普通的生理变色是体色深浅的改变,这在许多蟹中可以发现。而虾类常有更广泛的颜色变化,例如一种小长臂虾(Palaemonetes),色素细胞中含有红、黄、蓝三种色素颗粒,通过其中任一色素颗粒的独立移动使之可以适应任何颜色的背景。许多虾类都具有这种能力。色素颗粒在色素细胞中的移动受眼柄中分泌的激素(hormones)控制。例如许多虾的色素细胞含有红、黄、蓝和白色素,移走眼柄,红、黄色素扩散而使体色变暗。如注射眼柄的激素提取物,则白色素扩散而使体色迅速变浅。因此,对每种色素可能都存在着一对对抗性的促色素细胞素(chromatophorotrophins),其中一个处于分散状态,另一个处于集中状态。体色深浅的改变,取决于一对对抗性促色素细胞素中的哪一个处于扩散状态以及哪一个处于集中状态。