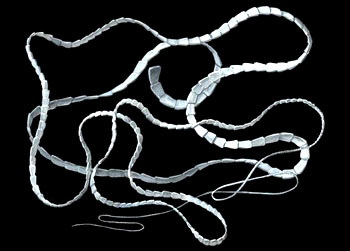
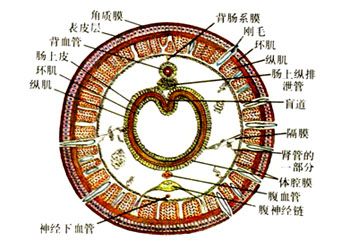
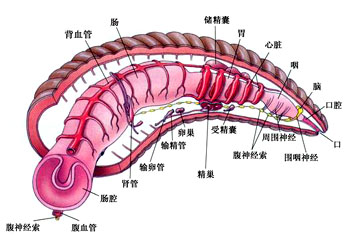
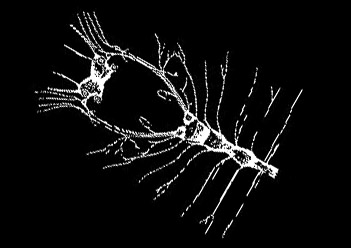
三叶虫亚门・ 三叶虫纲螯肢亚门・ 肢口纲・ 蛛形纲・ 海蜘蛛纲甲壳亚门・ 头虾纲・ 鳃足纲・ 软甲纲・ 颚足纲六足亚门・ 原尾纲・ 弹尾纲・ 双尾纲・ 昆虫纲多足亚门・ 倍足纲・ 唇足纲・ 少足纲・ 综合纲
|
三叶虫亚门・ 三叶虫纲螯肢亚门・ 肢口纲・ 蛛形纲・ 海蜘蛛纲甲壳亚门・ 头虾纲・ 鳃足纲・ 软甲纲・ 颚足纲六足亚门・ 原尾纲・ 弹尾纲・ 双尾纲・ 昆虫纲多足亚门・ 倍足纲・ 唇足纲・ 少足纲・ 综合纲
|
|||
你的位置: 首页> 动物界> 节肢动物门
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||