头索动物亚门
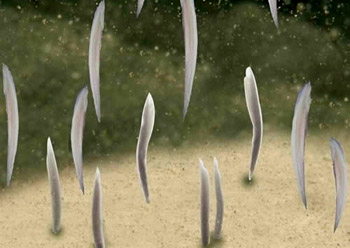
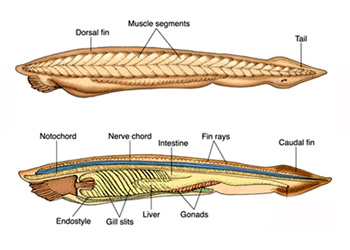
又称狭心纲(Leptocardii),仅文昌鱼目(Branchiostomiformes),包括文昌鱼科(鳃口科)(Branchiostomidae)和偏文昌鱼科(侧殖文昌鱼科)(Epiyonichthyidae)。文昌鱼科生殖腺左右对称排列于腹部两侧,有1属9种,包括白氏文昌鱼(Branchiostoma belcheri)、加州文昌鱼(Branchiostoma californiense)、开普斯文昌鱼(Branchiostoma capense)、卡氏文昌鱼(Branchiostoma caribaeum)、佛州文昌鱼(Branchiostoma floridae)、普通文昌鱼(Branchiostoma lanceolatum)、瓦氏文昌鱼(Branchiostoma valdiviae)、维吉尼文昌鱼(Branchiostoma virginiae)。其中加州文昌鱼产于北美圣地亚哥湾,长达100毫米,是体形最大者;我国最早发现的是厦门白氏文昌鱼(Branchiostoma belcheri),广泛分布于渤、黄、东、南海的浅水区。偏文昌鱼科仅右侧有一行生殖腺,有2属5种,包括偏文昌鱼属(Asymmetron)和侧殖文昌鱼属(Epigonichthys)。前者包括芦卡偏文昌鱼Asymmetron lucayanum和莫氏偏文昌鱼Asymmetron maldivense,后者包括短刀侧殖文昌鱼Epigonichthys cultellus、芦卡侧殖文昌鱼Epigonichthys lucayanus和马尔代夫侧殖文昌鱼Epigonichthys maldivensis。我国有偏文昌鱼(Asymmetron maldivense)和芦卡侧殖文昌鱼(Epigonichthys lucayanus),分布于台湾南端的南湾水域。