多毛纲
寡毛纲
蛭纲
你的位置:
首页
>
动物界
>
环节动物门
> 寡毛纲
寡毛纲Oligochaeta包括常见的蚯蚓、颤蚓等一类动物,一般认为寡毛类是由海产穴居的原始环节动物侵入淡水及陆地而发展起来的一支,它们以明显的特征独立于多毛类:身体分节但不分区,疣足退化,体表具刚毛,但刚毛的数目远远少于多毛类,因此称寡毛类。雌雄同体,生殖腺1~2对,有体腔管起源的生殖导管,性成熟时体表出现环带(clitellum),交配时可相互授精,卵产于环带中,脱落后形成卵茧,直接发育。约有3000种左右。寡毛类的分类方法过去一直是根据雄性生殖孔在精巢体节隔膜的前后而分为近孔目(Plesiopora)、前孔目(Prosopora)及后孔目(Opisthopora)三个目。近年来Jamieson(1978)根据生殖腺、环带及刚毛等结构将寡毛类分为带丝蚓目(Lumbriculida)、颤蚓目(Tubificida)和单向蚓目(Haplotaxida)。
寡毛纲
Oligochaeta
寡毛纲的主要特征
多为陆生,少数为淡水生
头部不明显,感官不发达
无疣足,刚毛直接生于体壁上
雌雄同体,具生殖环带,直接发育
带丝蚓目 Lumbriculida
仅带丝蚓科(Lumbriculidae)。每个体节具4对刚毛,精巢一对,雄性生殖孔就在精巢所在体节。卵巢1~2对,环带很薄,包括雄性生殖孔及雌性生殖孔,例如带丝蚓(Lumlbriculus),淡水生活。
颤蚓(Tubifex)
仙女虫(Nais)
带丝蚓(Lumbriculus variegatus)
颤蚓目 Tubificida
刚毛4束,每束多超过2根,常呈发状,精巢、卵巢各一对,位于相邻的两个体节内,雄性生殖孔位于精巢体节之前或之后的相邻体节上,环带薄,但略隆起,亦包括雄性生殖孔及雌性生殖孔,淡水、海水生活,个别的种陆地生活,可分为两个亚目。颤蚓亚目(Tubificina):受精囊孔在雄性生殖孔之前或之后的相邻体节上,很少在相同的体节上,刚毛多样,例如颤蚓(Tubifex)、水丝蚓(Limmodrilus),仙女虫(Nais)、尾盘虫(Dero)等。线蚓亚目(Enchytraeina):受精囊在精巢之前,两者相距五个体节,刚毛简单,如白丝蚓(Fridericia)。关于体虫类(Aeolosomatidae),这是一种淡水常见的小型种类,过去认为它们是原始的寡毛类,属于近孔目。但它们大量的体节可以产生配子,没有明确的生殖腺,也没有生殖导管,这些特征反与多毛类相似,因此Jamieson主张这类动物与寡毛类无关。所以有关其分类地位,尚有待进一步研究。
赤子爱胜蚓(Eisenia foetida)
陆正蚓(Lumbricus terrestris)
绿色异唇蚓(Allolobophora chlorotica)
环毛蚓(Pheretima)
陆正蚓(Lumbricus terrestris)
单向蚓(Haplotaxis gordioides)
单向蚓目 Haplotaxida
通常是两对精巢位于两个体节,随后为二对卵巢体节。但也有的种仅一对精巢,或仅一对卵巢,或两者均一对,如仅一对精巢,其卵巢必相隔1―2个体节。雄性生殖孔在精巢之后一或几个体节上。分为四个亚目,其中Alluroidina亚目为南美及非洲产。单向蚓亚目(Haplotaxina):具4束简单的或分叉的刚毛,每束2根。精巢在第10和第11节上,卵巢在第12节和13节上,或只在第12节上,雄性生殖孔在精巢之后一节,环带薄,淡水或半陆生生活,仅单向蚓科(Haplotaxidae),如单向蚓(Haplotaxis)。链胃蚓亚目(Moniligastrina):每节4对简单的刚毛,精巢1~2对,位于精巢囊中,雄性生殖孔1~2对,在相关精巢囊之后一节的后缘处,卵巢一对,环带薄,有的是大型蚯蚓,例如链胃蚓(Moniligaster)、杜拉蚓(Drawida)等。正蚓亚目(Lumbricina):刚毛简单,8个,有时很多,排成环状,精巢1~2对,一般在10~11节,雄性生殖孔一对,位于后精巢之后2个或更多的体节上,即在14节之后。卵巢一对,位于第13节,环带较厚,卵黄较少,主要陆生,少数为水生或半水生,包括大量常见的蚯蚓。如雄性生殖孔开口在第15节环带前的正蚓(Lumbricus)、爱胜蚓(Eisenia)、异唇蚓(Allolobophora);雄性生殖孔开口在第17节的寒蚓(Ocnerodrilus);雄性生殖孔开口在第18节的巨蚓(Megascolex)、环毛蚓(Pheretima)、微蠕蚓(Microscolex)等。赤子爱胜蚓(Eisenia foetida)、毛里巨蚓(Megascolex mauritii)(属巨蠕蚓科(Megascoleci-dae)和无锡微蠕蚓(Microscolex wuxiensis)(属棘蚓科Acanthodrilidae)为我国著名的发光蚯蚓。我国已应用分析染色体组型方法鉴定蚯蚓,赤子爱胜蚓的染色体2n=22,与人工养殖的蚯蚓“太平二号”和“北星二号”完全相同(许智芳1983)。
寡毛纲的生态与分布
寡毛类从生态上可区分成陆生及水生的两种类型。大量的种是陆生的,体型也较大,在陆地上穴居。除了沙漠地区,任何土壤中都有分布,例如各种蚯蚓。有人报导每平方米草地的土壤中可有8000条线蚓(Enchytraeid)和700条正蚓(Lumbricids)。它们主要在土壤的表层分布,那里有机质比较丰富。土壤的结构、酸碱度、含水量、通气性等都是限制其分布及数量的因素。例如土壤中的酸、碱度对寡毛类有很大的限制作用,酸性土壤不利于他们的生存,因土壤中缺乏游离的钙离子,而钙离子是维持其血液pH值的重要因素,所以酸性土壤中寡毛类较少。一些大型的种类在环境不利时,例如在干旱或寒冷时,可潜入土壤深层,有时达1m多。它们靠身体的头端不断地挖掘,吞噬土壤,并分泌粘液,做成穴道。另一类寡毛类是水生的,主要分布在各种淡水水域,特别是有机质丰富的浅水。一般水生种类体型较小,结构简化,有些特征与原环虫相似,但一般认为可能是结构的简化而不是原始性状。水生种类多在水域中的植物表面爬行,取食沉渣,也有的种在水底软泥或沉积物中穴居,还有少数种类在河口处生活。许多种类是世界性分布的。水域中寡毛类数量的多少常标志着水质污染的程度。
寡毛纲的形态与生理
蚓螈
属于
两栖纲
蚓螈目
,与蚯蚓极为相似,皮肤裸露,上有多数环状皱纹和粘液,无四肢及肢带;通常在湿地营洞穴生活,眼退化隐于皮下,耳无鼓膜 (图为粗线真蚓 Caecilia pachynema )
蚓蜥
属
爬行纲
蚓蜥目,体长圆柱形,具浅沟,无外耳,眼退化,均无后肢,多数无前肢,穴居,头顶具大型坚硬鳞片,用以钻洞;与蜥蜴目近缘(图为地中海蚓蜥Blanus cinereus)
盲蛇
属
爬行纲
蛇目
,小型蛇类,形似蚯蚓;眼小,不明显,隐于半透明的眼鳞下;背鳞、腹鳞分化不明显;掘土穴居(图为钩盲蛇Ramphotyphlops braminus)
外部形态
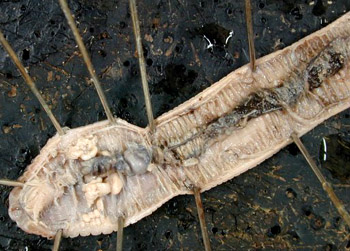
:寡毛类体型大小差别很大。最小的个体不足1mm,如原口虫科(Aeolosomatidae)。最大的蚯蚓体长达1~3m,身体通常圆柱形,有时略扁,体表分节明显。陆生的蚯蚓体节数较多,最多的可达600节,一般在100~200节之间。水生的种类不仅体小,体节数也少,一般具6~7节或十几节不等。口前叶不发达,常成一小叶状或锥状盖在口上,口前叶也不具触手、触须等结构。如仙女虫(Nais),口均位于围口节上,咽不外翻,咽上无齿,肛门开口在身体末端。寡毛类没有疣足,但体表有刚毛,刚毛的数目、排列方式在不同种有所不同。水生种类刚毛较长,陆生种类较短。一般每个体节有一对侧刚毛束(或背侧刚毛束)及一对腹刚毛束,它们代表着多毛类疣足的背、腹叶遗迹。每束刚毛的数目为1~25不等。大多数陆生及水生种类刚毛的数目是8根,成4束,每束2根,这种排列称对生刚毛(lumbrieine seta),例如正蚓。也有的种刚毛数很多,每节几十个环绕体节分布,这种排列称环生刚毛(perichaetineseta),例如环毛蚓(Pheretima)。刚毛的形状因种而异,可形成毛状、钩状、叉状、S状等多种。刚毛是由体壁中表皮细胞形成的刚毛囊分泌的。刚毛囊有伸肌及缩肌控制其运动,每个刚毛囊可分泌一根或一束刚毛,刚毛脱落后可重新分泌形成。体表除了有口与肛门分列于身体两端之外,还有肾孔、生殖孔开口在固定的体节上。陆生的种类在背中线上还有数目不等的背孔(dorsal pores),它是体腔直接与外界相通的小孔。由小孔排出的体液可以滑润及潮湿皮肤,有助于在土壤中钻行。性成熟时,身体前端的几个体节体壁的腺体加厚,膨胀而形成一环带(Clitellum),这是寡毛类动物的第二性征,即性成熟时才出现的特征。水生种类环带仅出现在1~2个体节内。环毛蚓是由3个体节组成,位于第14~16节内,其中含有许多细胞。细胞能分泌粘液,交配后的粘液形成卵茧。另外细胞分泌的类蛋白物质(albuminoid material)有滋养胚胎的作用。
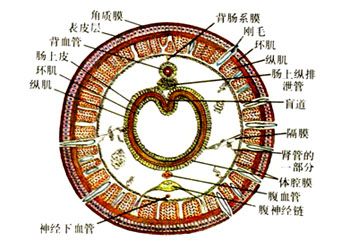
体壁、运动及体腔
:寡毛类,特别是陆生种类,体壁的结构与穴居的多毛类相似,即由角质层、表皮细胞、环肌及纵肌组成,肌肉层内为体腔膜。表皮细胞中有发达的腺细胞,特别是陆生种类,可以分泌粘液湿润皮肤,以利于在土壤中运动。水生种类肌肉层较薄。寡毛类的运动方式为蠕动收缩,相似于钻穴的多毛类。几个体节成为一组,一组内纵肌收缩,环肌舒张,体节则缩短,同时体腔内压力增高,刚毛伸出以附着。而相邻的体节组环肌收缩,纵肌舒张,体节延长,体腔内压力降低,缩回刚毛。每个体节组与相邻的体节组交替收缩纵肌与环肌,使身体呈波浪状蠕动前进。蚯蚓每收缩一次可前进2~3cm,收缩方向可以反转,因此可做倒退运动。寡毛类体腔发达,同时还被发达的隔膜分割成按节排列的体腔室。隔膜上有小孔及括约肌,以控制体腔液由一个体节流入另一个体节。特别是陆生种类,隔膜肌比水生种类发达。体壁肌肉收缩时,隔膜肌可以调节体腔内的压力,协助体节的延伸。身体背中线节间沟处有背孔一个,排出的体腔液可使体表湿润。体腔中还包含有体腔细胞、蛋白质及悬浮的其他颗粒。
蚯蚓横截面结构
寡毛类疣足退化,刚毛直接长在身体上
寡毛类的运动方式为蠕动收缩
取食与营养
:寡毛类中一些小型的淡水种类可以取食沉淀物、藻类或原生动物。它们以身体前端的刚毛收集食物,由刚毛送入口中,例如仙女虫。绝大多数的种类是腐食性的,它们以腐烂的植物或其他有机物为食。陆生的种类吞食土壤及砂粒,以获取其中的有机物质,例如蚯蚓。寡毛类的消化道为一直管,口位于围口节上,口后为一很小的口腔,口腔后为咽。水生种类咽可伸出或外翻,陆生种类咽不能伸出。咽壁具发达的肌肉,形成球状,有泵的抽吸作用。蚯蚓及管盘虫(Aulophorus)等的咽壁上还有大量的肌肉纤维连接到体壁上,以至形成一肌肉质盘,包在咽的周围,以增强其抽吸作用。咽壁内有发达的单细胞腺体,它所分泌的粘液可使食物颗粒粘结,其中含有蛋白酶可对食物进行初步消化。有的种类咽发生几度紧缩,以致形成几个咽球。咽后为窄长的管状食道。陆生种类的一个特征是食道上有一对或几对钙腺(calciferous glands)位于食道两侧,是由食道壁内陷形成的一种腺体,它可分泌钙质,以减少体内随食物进入的过多的钙,并通过控制离子的浓度以维持体液与血液的酸碱平衡。水生种类不存在钙腺。食道之后形成嗉囊(crop)及砂囊(gizzard),前者是一薄壁的囊,用作食物的暂时贮存,有的种类嗉囊不发达或缺乏。后者是一厚壁的囊,内表面有一层厚的几丁质层,用以研磨食物成为细粒。砂囊后为管状的胃,胃上有丰富的血管及腺体。胃后为肠,它纵贯其余体节,是其消化及吸收的主要场所。肠的前端上皮细胞能分泌蛋白酶和纤维素酶等以进行食物的消化。消化后的营养物质由血液送到全身。蚯蚓等陆生寡毛类在肠的前端还形成一对发达的盲肠,环毛蚓盲肠在第26节处,盲肠内有发达的腺体,也是消化的重要场所。肠道中段背中线处内陷形成盲道(typhlosole),以扩大消化及吸收面积。肠壁的外周体腔膜细胞改变成黄色细胞,或称黄色组织(chloragogen tissue),它在物质的中间代谢中起重要作用,即它是脂肪及糖元合成及贮存的中心,同时也能使蛋白质脱氨基而分解成氨及尿,有一定的排泄作用。黄色组织是由于细胞内黄绿色脂类内含物的存在而成为黄色,这些细胞也可以转变成油细胞(eleocytes)而被释放到体腔液中。肠的后端少数体节为废物的贮存处,内含蚓粪,最后由肛门排出体外。
呼吸
:寡毛类是通过体表的气体扩散而进行呼吸作用。一些小型的水生种类,例如尾盘虫(Dero)在身体的后端有一圈指状或丝状的突起,起着鳃的作用。管盘虫(Aulophorus)、仙女虫是以外界的水通过肛门不断地流入及流出直肠而进行呼吸,一般它们没有呼吸色素,气体直接进行扩散。陆生的寡毛类也没有呼吸器官,而是由于体表分布有大量的微血管网,在皮肤潮湿的情况下,很容易的进行气体的交换。蚯蚓的血浆中溶解有丰富的血红蛋白,血红蛋白很容易与氧结合及释放出氧。有人报导正蚓所利用氧的40%是由血浆中的血红蛋白进行传递的。也有一些水生的寡毛类可以在低氧的条件下生存,例如颤蚓(Tubifex)。这是一种血浆中溶有血红蛋白的小型寡毛类,身体呈红色,常在池塘底部淤泥中营管居生活,那里氧气含量极少,但血浆中的血红蛋白很容易与氧结合而进行氧的传递,甚至在无氧的条件下也能使其生存一段时间。为了更易于获得氧,它们常将身体的后端伸出淤泥之外,以利于更有效地进行呼吸作用。
循环
:寡毛类的循环系统与多毛类相似。原始的种类仅有背血管、腹血管及连接两者的少数几对心脏,例如体虫。大多数较进化的种类,血管系统的结构也较复杂,例如环毛蚓在消化道的背面有一条背血管。由于血管管壁的肌肉较发达,管内尚有瓣膜,靠其波状收缩,迫使血液由后向前流。背血管及心脏决定着血液的流向,背血管中的血液流到身体前端后,一部分血液分布到食道、咽、脑等处,大部分血液经过4对心脏流入腹血管。心脏中也有瓣膜,可以有节奏地跳动,也起控制血液流向的作用。腹血管位于消化道腹面,不能搏动,腹血管中的血液由前向后流,腹血管在每个体节都有血管分支,分布到体壁、肠道及肾管等处,在那里形成微血管网。经过气体及物质的交换之后,前14节的血液流入消化道两侧的食道侧血管(lateral esophageal vessel),14节之后经交换后的血液流入腹神经之下的神经下血管(subneural vessel)。食道侧血管与神经下血管是相连的,血液也是由前向后流。神经下血管中的血液再通过每节一对的壁血管(parietalvessel)流回背血管,背血管也接受肠血管(intestinal vessel)的血液。如此循环,完成物质的传递功能。寡毛类的循环系统在不同的种可能有所改变,例如正蚓具有五对心脏,没有食道侧血管,而有神经侧血管。颤蚓仅有一对心脏。许多种类血液无色;少数种红血蛋白存在于血细胞中。
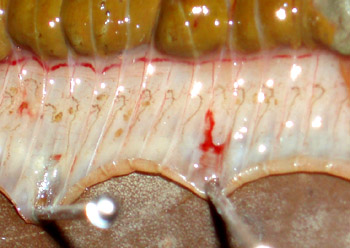
寡毛类每体节都有一对后肾管(图中每体节褐色部分)
蚯蚓将卵产在卵茧中,卵在其中受精(图中黄色部分)
卵茧呈卵圆形,淡黄色,其中有卵粒1~20不等
排泄
:成年的寡毛类除了两端几个体节之外,其余每个体节都有一对后肾为其排泄器官。后肾的结构相似于多毛类,具漏斗状带纤毛的肾口。肾管很长,每节的肾管穿过体节后端的隔膜后盘旋,在肾管周围有腹血管分出的血管网包围,肾管的后端变粗形成膀胱。后肾主要通过肾口在体腔中收集代谢产物,同时由于血管网的包围也能主动收集来自血液中的代谢产物,回收有用的盐离子及水分。水生的寡毛类其排泄物主要是氨,陆生种类是氨和尿素,最后形成比体腔液及血液低渗的尿,在每个体节腹侧面经肾孔排出体外。后肾对水分及某些盐类有重吸收作用。但环毛蚓没有这种每节一对的典型后肾,而是在每个体节内有为数极多的小肾管,其数目可达数百个。每个小肾管的构造与大肾管相似,它们分布在体壁的内表面、隔膜的前后壁以及咽头食道上。其中体壁小肾管开口于体外,隔膜及咽头小肾管开孔到肠道,因此其排泄物经肠道肛门而排出体外。排泄物经过肠道时,其中的水分被吸收,因此有利于它们在干燥条件下生存。有些蚯蚓在干燥条件下可钻入土下3米深处,停止活动,即使体内水分丧失70%也不致死亡。而一旦解除干燥,又能很快地恢复活动。
神经与感官
:寡毛类具典型的链状神经系统。结构简单的种类如体虫(Aeolosoma),它的脑位于原口节,脑与腹神经索均位于上皮内,没有咽下神经节。而大多数的寡毛类神经结构脱离上皮,而位于体壁肌肉层之内,即体腔内。脑位于第三体节咽的背面,由脑发出神经到口前叶及口腔等,在围咽神经环及腹神经索连接处形成咽下神经节,由它发出神经到前端体壁上。咽下神经节是其运动及反射的控制中心,并控制整个腹神经索,脑仅控制身体的协调。实验证明摘除脑,其运动不受影响,但不能协调改变了的环境。如破坏咽下神经节,则所有的运动都停止。咽下神经节之后为神经链,每体节的神经节分出三对神经到体壁、内脏肠道等处,其中包括感觉纤维及运动纤维。蚯蚓具有简单的反射弧(reflex arc),它包括三种神经元:(1)感觉神经元(sensory reuron),其细胞体位于体壁表皮细胞中,它感受刺激后经神经纤维(也称传入纤维)到达中枢(神经节内)。(2)联络神经元(association neuron),其整个细胞均在神经节内,它接受感觉神经传入的冲动,再传递到运动神经元。(3)运动神经元(motor neuron),细胞体位于中枢内,其神经纤维(也称传出纤维)传出冲动到效应器,如肌肉、腺体等。各种神经元之间不直接接触,而是通过突触传递(synapsis),这种感觉细胞感受刺激使效应器产生反应,如肌肉收缩或腺体分泌就是一个反射。一个体节的收缩可通过反射作用引起相邻体节的收缩,以致形成部分体节的收缩波。大多数的寡毛类在神经索中有巨大神经纤维,一般5条,其中3条显著,位于神经索的中背部。中间的一条巨大神经向尾端传导冲动,两侧的两个巨大神经纤维向头端传导冲动。另两条不显著,彼此分离,位于神经索的中腹部。巨大神经传导冲动的速度数倍或十几倍于普通神经,因此当身体的任何一点受到刺激,通过巨大神经纤维的传导都可引起所有体节同时收缩,以迅速逃避或隐藏于穴中。有报导证明,正蚓的脑中发现有神经分泌细胞(neurosecretory cell),它所产生的分泌物具有激素(hormone)的性质,能调节身体水与盐分的平衡,也能调节生殖活动。一些水生的寡毛类具有眼。眼呈色素杯状,结构简单,是它们的主要感觉器官。陆生种类由于在土壤中钻穴生活,感官不发达,无眼,其感觉功能主要是由分散于表皮中的感觉细胞来完成。例如一些感觉细胞聚集在一起,在皮肤表面形成小的突起(图9-39A),或成堆的感觉细胞形成一结节,并伸出长的突起到体表,这种感受器可能有触觉及化学感觉的功能。这些结构主要分布在身体的前、后端及腹面两侧。表皮内还有独立的光感受细胞,呈晶体状,具突起进入上皮下,并与脑神经分支相连,它对光的强弱有反应,主要分布在头、尾两端的背面,所以蚯蚓趋向弱光,回避强光的本领是与光感受细胞联系在一起的。此外,体壁上还分布有丰富的神经末稍,也有触觉功能。
生殖与发育
:寡毛类主要行有性生殖,但一些水生种类无性生殖也相当普遍,例如管盘虫、仙女虫等。有实验报导管盘虫在三年内无性繁殖150代,而没出现过有性生殖。其无性生殖方式是以横分裂为主,即先在某个体节处出现了新细胞形成的分裂带,以后在分裂带处断裂。也有的种是以出芽方式进行的。水生寡毛类也具有较强的再生能力。绝大多数种类为雌雄同体。少数水生种类缺乏明确的生殖腺,即许多体节均可形成生殖细胞,也没有生殖导管,而是由肾管排出精、卵,如体虫(Aeolosoma)。陆生种类有明确的生殖腺,且仅限于有限的几个体节之内。一般具有二对精巢,1~2对卵巢,精巢在前,卵巢在后,具单独的生殖导管,它们的数目及位置是分类的依据之一。生殖细胞来自体腔隔膜上的上皮细胞,例如环毛蚓具有二对精巢囊,分别位于第10,11体节内,每对精巢囊的后方各有一对由体腔隔膜形成的贮精囊,位于第11,12体节内,并与精巢囊有小孔相通。当精细胞在精巢囊中形成后,需在贮精囊中发育成熟,然后再回到精巢囊中,经其中的精漏斗进入输精管。每侧两条输精管紧密并行,穿过数个体节后至第18节与前列腺管汇合,并由雄性生殖孔开口腹面两侧。生殖孔的周围有前列腺,它的分泌物有滋养精子和帮助交配的作用。雌性生殖系统包括一对卵巢,位于第13节,经卵漏斗进入很短的输卵管,以共同的雌性生殖孔开口在第14节的腹中线上。与雌性生殖系统相关的还有2~3对受精囊(spermatheca),位于第6~9节内,每一囊在后一体节腹面节间沟处单独开孔,用以贮存交配后的精子。寡毛类与生殖相关的结构还有环带,它与性成熟相关,仅在性成熟时出现。它是由相邻的几个体节体壁的上皮细胞膨大、并分布有大量的单细胞腺体所形成。例如环毛蚓包括三个体节(第14~16节),水生寡毛类由二个体节形成,正蚓有6~7个体节形成,最多的种可达60个体节。蚯蚓的环带中有粘液腺(mucousgland)、卵茧分泌腺(cocoon secreting gland)及白蛋白腺(albumingland),它们的分泌物分别具有协助交配、形成卵茧、分泌白蛋白使卵悬浮于卵茧中,并获得营养等功能。寡毛类虽为雌雄同体,但仍需交配受精。交配时两个虫体的前端腹面以头、尾相反的方向相互吻合,大多数是以一方个体的雄性生殖孔对准另一方个体的受精囊孔,这时环带可分泌粘液使虫体连接在一起,体表的刚毛也使虫体相连。然后相互赠送精子,每个受精囊装满对方的精子约需1.5小时。正蚓的交配不同于上述的环毛蚓,因为它的环带是在雄性生殖孔之后而不是雄性生殖孔之前,所以交配时是以环带对准对方的受精囊孔,精子排出后,沿着环带分泌的粘液沟游动一定距离后再到达受精囊孔,经2~3小时后完成授精,随后两个个体各自分开。交配数日后,卵开始成熟。这时环带向外分泌粘液,并包围环带,凝固后形成茧管,环带再分泌白蛋白,成熟的卵排入其中。这时环带与茧管彼此分离,随着虫体的蠕动,茧管逐渐前移,当移到受精囊孔所在体节时,精子由该孔注入其中。当茧管完全脱离虫体后,其外层的粘液物分解将两端封闭,形成卵茧,卵在其中受精。卵茧呈卵圆形,淡黄色,其中有卵粒1~20不等。在生殖季节内数日后即可产生一个卵茧,连续不断可形成多个。一般陆生种类的卵较水生种类的卵小,卵黄也少。受精卵在茧中发育,茧中的白蛋白为胚胎提供丰富的营养。蚯蚓均直接发育,受精卵经不等的完全卵裂形成中空的囊胚,经内陷法形成原肠胚,由端细胞形成中胚层带,裂腔法形成体腔。发育进行的时间因种而异,由一周到数月不等。环毛蚓需2~3周,正蚓需12~13周。寡毛类动物的寿命由一年到数年,一般水生种类的寿命较短,陆生种类寿命较长。正蚓有的可生活长达6年,一般一年后性成熟。颤蚓每年仅繁殖一次,随后其生殖系统被吸收,来年再重新形成。
蚯蚓吞食土壤和有机物质的能力很大,可利用蚯蚓处理城市的有机垃圾,保护环境,防止污染,可化害为利,抑制公害
陆蚓类穴居土壤中,在土壤中穿行,吞食土壤,使土壤疏松,改良土壤的理化性质,增加腐殖质,对土壤团粒结构的形成起很大作用
蚯蚓含蛋白质较高,其含量约占干重的50%~65%,是一种动物性蛋白添加饲料,对家禽、家畜、鱼类的产量提高效果明显
蚯蚓在改良土壤中的作用
如果说把单位体积内水生寡毛类的数量作为水质污染程度的一种指标,那么,单位体积内陆生寡毛类的数量也可作为土壤肥力的一种指标。蚯蚓对土壤的改良有着重要的作用,这表现在:(1)粉碎及分解有机物蚯蚓对落入土壤中的植物有机物的分解起着重要作用。由于它们的取食习性,大量的表面落叶层的植物被消化及分解,使表面形成一层肥沃的未被完全分解的有机物碎屑,形成土壤疏松的表层,为其他生物的进一步分解创造了条件。这对果园、森林等落叶层地带尤为重要。(2)促进土壤微生物的作用进入土壤中的有机物经蚯蚓的初步分解后,将由土壤中的微生物做进一步地分解。有人报导在正蚓的消化道中分离出50余种微生物,其种类与土壤中的群落是一致的。蚓粪也是土壤微生物传播的一种方式,土壤中的蚯蚓可增加土壤微生物数量的5~10倍,这加速了土壤中有机物的进一步分解及土壤的腐植化过程。(3)对土壤的翻动及团聚作用由于蚯蚓的钻穴习性可以将表层含有机物丰富的土壤与底层有机物含量很少的土壤加以搓合,其搬动量随蚯蚓的数量及土质可达2~250吨/年,使翻到表层的土壤达0.1~5cm厚,这样也增加了土壤的通气性,使土壤中空气的含量由8%提高到20~30%。土壤的毛细作用大大增加,提高了对降雨量的吸收及保持。另外,蚯蚓肠道中腐败的有机物与钙腺中的钙形成腐质酸钙,将土壤颗粒粘合成团聚体,土壤中的微生物也参与了这一过程,使土壤的团聚结构得到改善。(4)增加土壤肥力土壤中大量的蚯蚓不断地繁殖与更替,腐烂的尸体使土壤中氮的总含量得到增加。有实验证明,在12℃时蚯蚓尸体2~3周内完全解体并分解,其中氮的25%形成硝酸盐,45%形成氨。如果按每条蚯蚓产生10mg的硝酸盐计算,如每公顷有300万条蚯蚓,那么一公顷土壤即可自然增加217公斤的硝酸盐肥料。如果蚯蚓的寿命平均按一年计算,那么它所产生的氮量也是相当可观的。
水蚓类都可作为淡水鱼类的饵料,如颤蚓
蚯蚓体内含地龙素、多种氨基酸、维生素等,自古即入药
蚯蚓制成的地龙粒肥
寡毛类动物的经济意义
寡毛类中的水蚓类都可作为淡水鱼类的饵料,但它们繁殖过多时,可损害鱼苗或堵塞输水管道。陆蚓类穴居土壤中,在土壤中穿行,吞食土壤,能使土壤疏松,改良土壤的物理化学性质。经过蚯蚓消化管的土壤,排出成蚓粪,含有的氮、磷、钾的成分较一般土壤高数倍,是一种高效有机肥料。蚓粪又可增加腐殖质,对土壤团粒结构的形成起很大作用,有人估计林地或果园每年由蚯蚓形成的土壤团粒结构每公顷达47t~170t,增加氮素75kg~125kg。同时蚯蚓还可将酸性或碱性土壤转化为近中性。蚯蚓含蛋白质较高,其含量约占干重的50%~65%,含18~20种氨基酸,其中10余种为禽畜必需的。故蚯蚓是一种动物性蛋白添加饲料,对家禽、家畜、鱼类的产量提高效果明显。蚯蚓体内含地龙素、多种氨基酸、维生素等,有解热、镇静、平喘、降压、利尿等功能,自古即入药。蚯蚓吞食土壤和有机物质的能力很大,可利用蚯蚓处理城市的有机垃圾,保护环境,防止污染,可化害为利,抑制公害。各国都兴建养殖蚯蚓的工厂,繁殖蚯蚓,处理废料,生产有机肥料。蚯蚓又有聚集土壤中某些重金属的能力(镉、铅、锌等),可收集蚯蚓处理受重金属污染的土壤,达到减轻污染的目的。蚯蚓加工后可制作食品,国外有利用蚯蚓制饼干、面包等。