海星纲
砂海星(Luidia ciliaris)
砂海星(Luidia foliolata)
砂海星(Luidia australiae)
指海星(Linckia laevigata)
珠海星(Fromia monilis)
槭海星(Astropectinidae)
光海星(Psilaster charcoti)
网瘤海星(Oreaster reticulatus)
面包海星(Culcita novaeguineae)
鸡爪海星(Henricia)
驼海燕(Asterina gibbosa)
拟海燕(Patiriella parvivipara)是最小的海星,直径仅4毫米
太阳海星(Solaster)
长棘海星(Acanthaster planci)
墨西哥海星(Plectaster decanus)
棘轮海星(Crossaster papposus)
红海盘车(Asterias rubens)
多棘海盘车(Asterias amurensis)
多棘海盘车(Asterias amurensis)群体
菊海星(Heliaster)
多腕葵花海星(Pycnopodia helianthoides)
冠海星(Stephanasterias albula)
翼海星(Pterasteridae)
短刺豆海星(Pisaster brevispinus)
巨豆海星(Pisaster giganteus)
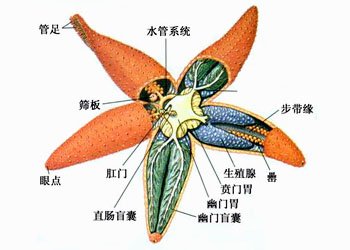
海星体呈五角星状,由体盘及腕组成,二者界限不明显;腕一般5条,故体呈典型的五辐射对称
海星各腕腹侧中央有一条自口伸向腕端部的步带沟,其内伸出管足2排,管足的末端有吸盘
海星的管足为运动器官,末端有吸盘,由于管足伸出向后撑,推身体前进;同时吸盘还有捕食功能
海盘车体表粗糙,许多内骨骼向外突出成棘或刺,棘间分布有叉棘和皮鳃,图为海盘车(Asterias rubens)
皮鳃呈泡状,为体壁自骨片间隙外凸形成。外层为上皮,内为体腔膜,体腔液在其内循环,有呼吸及排泄的功能
长棘海星(Acanthaster planci)表皮有极发达的刺,由内骨骼向外突出形成,图中白色突起即为皮鳃
水管系统是棘皮动物所特有的一个管状系统,来自次生体腔
水管系统由筛板、石管、环水管、辐射管、侧水管、管足等组成
水管系统的主要功能是运动
大多数海星类具长可弯曲的腕,管足上具吸盘,多以双壳类为食,取食时身体位于贝壳上,以两腕在贝壳两侧吸着,由于管足末端吸盘的真空作用,其拉力足以拉开双壳类的壳口
拉开壳口后,海星立刻翻出喷门胃插入壳口内,并分泌消化酶,直到闭壳肌及内脏部分地被消化,贝壳完全张开,再用胃包围吞彩澄镆煌进入口内;食物在胃内进行胞外消化,幽门盲囊中胞内消化
一些具短腕、管足上无吸盘的种类,是以较小的动物如小的甲壳类等为食,取食时是整个将食物吞玻消化在胃内而不在体外进行;深海生活的种类以纤毛过滤取食
海星为雌雄异体,精卵在海水中受精,经完全均等卵裂,有腔囊胚,以内陷法形成原肠胚。以体腔囊法形成中胚层和体腔,这与其他的无脊椎动物完全不同
椭圆形的幼体产生腕,体表纤毛退化,只在腕周围形成纤毛带,此为羽腕幼虫(图);后在背面基部产生二突起,进入短腕幼虫期,体两侧对称,前端具有3个小腕及一个吸盘
短腕幼虫在海水中游泳一时期后,沉入水底,营固着生活,进入变态期;逐渐由两侧对称变成辐射对称,原来胚的左侧成为身体的口面,右侧成为反口面;变态结束后发育为轴射对称的小海星