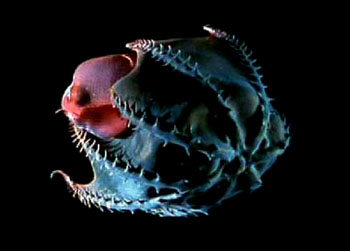
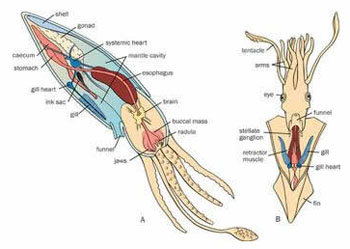
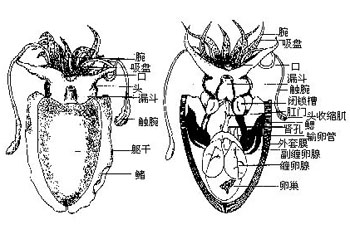
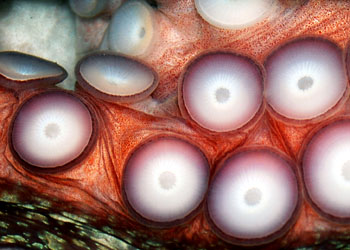
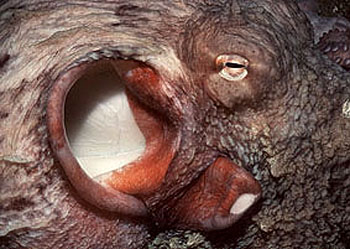
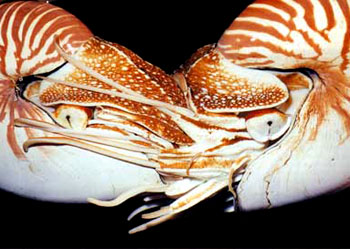
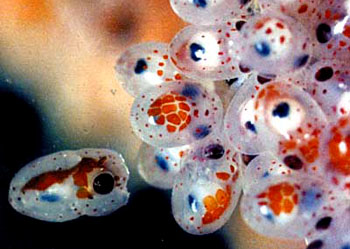
头足类的外套膜是多肌肉质的,其中包括放射肌与环肌。外套膜在身体的背面与体壁相连、腹面游离,与内脏之间的空间形成外套腔。由足特化形成的漏斗位于身体腹面躯干的前端,它也是肌肉质结构,漏斗的前端细长,其开口指向前端,漏斗后端宽大,可伸入外套腔中,漏斗后端两侧有一软骨凹陷与外套膜腹缘前端的软骨突形成一闭锁器(socket and knob)以封闭外套腔的开口。头足类的运动是以外套膜的肌肉收缩为动力。当外套膜环肌松弛、放射肌收缩时,外套腔体积扩大,水由外套腔开口处进入外套腔中,外套腔充水后,放射肌松弛、环肌收缩、闭锁器扣合关闭了外套腔的开口,外套腔中增加了压力,迫使水由漏斗前端开口处喷射出去。其反作用力推动身体迅速倒退,漏斗中有一前端游离的舌状活瓣,使水只能由漏斗口喷出,不能由外界流入。如果漏斗前端向后弯曲喷水时,则其反作用力推动身体向前运动。乌贼等动物在正常情况下漏斗向前喷水比向后喷水力量更大,因此头足类向后倒退运动较向前运动更迅速而常见。特别是在长距离游动时更常用倒退式运动。外套膜在躯干的边缘形成鳍,运动时起着舵的作用,加之许多种类身体又形成流线形,使它们的运动速度达到了水生无脊椎动物中最快的一类。