分类表
古多齿亚纲 隐齿亚纲 翼形亚纲 古异齿亚纲 异齿亚纲 异韧带亚纲
单板纲 无板纲 多板纲 腹足纲 掘足纲 瓣鳃纲 头足纲
双壳纲
古多齿亚纲(Palaeotaxodonta)两壳相等,能够完全闭合。贝壳表面具有黄绿色壳皮。壳内面多具有珍珠光泽。铰合齿数量多,沿前、后背缘分布。通常具有内、外韧带。前、后闭壳肌相等。鳃呈羽状。成体没有足丝。仅胡桃蛤目(Nuculoida)。又称湾锦蛤目,包括2总科
隐齿亚纲(Cryptodonta) 多等壳,小而薄,文石质。铰齿不发育,或具栉齿。外韧带。闭壳肌多为等柱,外套线完整,偶具小外套湾。现生代表具原鳃。海生,多内栖滤食。种类较少,仅1目1科,蛏螂目(Solemyoida),蛏螂科(Solemyidae),如蛏海螂(Solemya),我国不产
翼形亚纲(Pterimorphia)壳呈卵形、长方形或圆形,两壳相等或不等。壳顶两侧常具翼状的前、后耳。铰合齿多或退化,前闭壳肌较小或完全消失。多数种类具足丝,无水管,鳃为丝鳃型。包括4目。泥蚶、毛蚶、贻贝、江瑶、珍珠贝、扇贝、牡蛎等重要的经济贝类均属此亚纲
古异齿亚纲(Palaeoheterodonta)铰合齿分裂,或者分成位于壳顶的拟主齿和向后方延伸的长侧齿,或者退化。一般具有前、后闭壳肌痕各1个,两者大小接近。鳃构造复杂,鳃丝间和鳃瓣间以血管相连。仅蚌目(Unionoida)。背角无齿蚌(Anodonta woodiana)俗称河蚌,在我国各地江河、湖沼中均常见
异齿亚纲(Heterodonta)贝壳变化大,形状多样。铰合齿少,或者不存在。一般有前、后闭壳肌各1个,两者大小接近。鳃的构造复杂,鳃丝间和鳃瓣间有血管相连。外套膜通常有1~3个愈着点,在水流的出入孔处经常形成水管。包括帘蛤目(Veneroida)和海螂目(Myoida)
异韧带亚纲(Heterodonta)两壳经常不相等,壳内面一般具有珍珠光泽。铰合齿缺乏或比较弱。韧带常在壳顶内方的匙状槽中,而且常常具有石灰质小片。一般雌雄同体。包括笋螂目(Pholadomyoida)和隔鳃目(Septibranchida)。代表种如渤海鸭嘴蛤、菲律宾盘筒蛎、华贵杓蛤
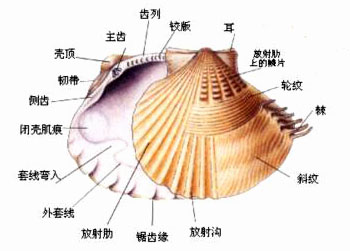
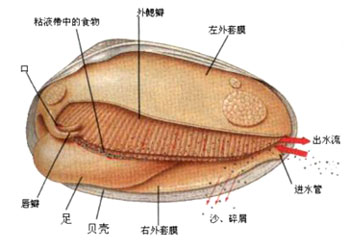
贝类外部结构模式图
最小的贝壳仅有2mm长,如珠蚬类(Sphaeriidae)
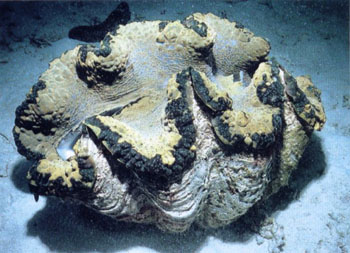
最大的贝壳长可超过1m,壳重达300kg,如砗磲(Tridacna)
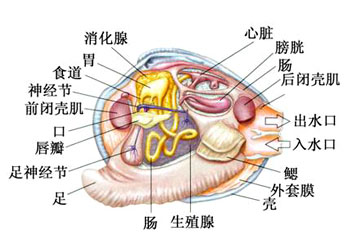
贝类内部结构图
蛤仔的出水口与入水口
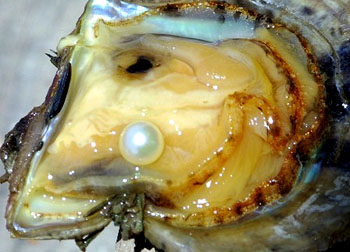
珍珠贝等的外套膜可将砂粒等异物包裹并分泌珍珠层,形成珍珠
较原始的种类多在浅层泥沙中穴居,例如原始的湾锦蛤、云母蛤(Yoldia),没有水管,它们在泥沙中运动时是通过足的两侧边缘的褶起与伸长
大多数双壳纲类生活在较深层的水中,它们的足变成斧状,它们的运动是由足部肌肉的收缩以及体内血压的改变而进行,有的种甚至可潜入泥沙中深达一米多,图为河蚬(Corbicula fluminea)
扇贝(Pecten)等自由生活的种类通过发达的后闭壳肌中横纹肌不停的收缩,而使两壳相互拍打,造成外套腔中水流迅速喷出而使身体在水中游动
大多数营固着生活的双壳类是用足丝固着,如贻贝(Mytilus)
牡蛎(Ostrea)是以左壳固着在岩石或海底木桩上
拟猿头蛤(Pseudochama)是以右壳固着在岩石表面
开腹蛤(Gastrochaena)等凿穴生活种类的壳的前端表面具锯状切齿,形成一个磨擦面,利用机械运动钻穴 ,根据不同的种或是由后缩足肌的收缩、壳的前端向上或下运动而凿穴
石蛏(Lithophaga)的外套膜可以分泌一种酸性粘液软化珊瑚骨骼或其他钙质贝壳而形成穴道,而其自身的钙质壳由于表面有角质层的保护而不受影响
船蛆(Teredo)以木质为食,消化道内有专门的消化腺可以分泌纤维素酶,以消化木质,许多的船舶由于船蛆的为害被蛀成空架而报废
淡水蚌的摄食机制
大多数贝类采取过滤取食方式
面盘幼虫是一些海洋贝类继担轮幼虫后的幼虫,具有游泳的面盘
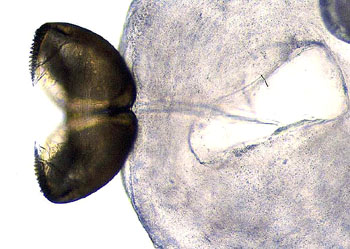
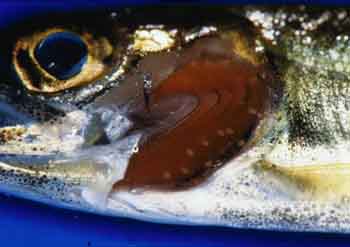
钩介幼虫是河蚌等特有的幼虫形态,长0.5~5mm长,具有双壳,壳的游离端具有钩与齿,壳内有足的原基,末端有粘着的足丝
属于鲤科,生殖时期的雌鱼出现产卵管,卵由产卵管直接产在蚌体内,并在其中受精发育;钩介幼虫同时也寄生在Q体表
钩介幼虫利用壳上的钩与齿或附着足附着在鱼鳃或皮肤上,寄主的被附着部位因受刺激而迅速分泌粘液形成包囊,幼虫在其中发育