分类表
前鳃亚纲 后鳃亚纲 肺螺亚纲
单板纲 无板纲 多板纲 腹足纲 掘足纲 瓣鳃纲 头足纲
腹足纲
前鳃亚纲(Prosobranchia)是腹足纲中最大的一个亚纲,生存种超过55000种。外套腔位于身体前端,鳃1~2个位于心耳之前,壳通常有板、侧脏神经索扭成"8"字形,因此又称为“扭神经类”。大多数雌雄异体,海水、淡水生活,极少数种可以陆地生活,包括原始腹足目、中腹足目和新腹足目3个目。
后鳃亚纲(Opisthobranchia)现存约1000种。具一个单栉鳃,一个心耳、一个肾。鳃位于心耳之后,壳与外套腔逐渐减少或完全退化消失。许多种本鳃消失,而出现次生性的皮肤鳃,身体又出现两侧对称。大多数种头部具2对触手,雌雄同体,全部海产。包括侧腔目、翼足目、囊舌目和裸鳃目4个目。
肺螺亚纲(Pulmonata)现存约20000种。本鳃消失,代之以右侧外套腔内壁充血形成“肺”进行气体交换,具一个心耳,一个肾;1~2对触角,触角端部或基部有1对眼;神经系统集中在前端食道周围。壳存在或成遗迹,成年后无厣板。雌雄同体,直接发育,陆生或水生。包括基眼目和柄眼目两个目。
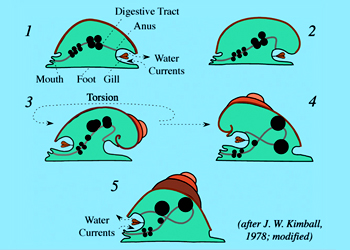
人们推测腹足类动物也是由原软体动物一样的祖先进化而来,首先这种祖先动物由于体积的增加及头、足经常缩入壳下,使内脏囊得到了充分的发展并不断地在身体背部隆起,结果由内脏囊顶端悬垂下来的外套膜及由外套膜分泌的贝壳都随内脏囊的隆起而增加高度,使身体背部及贝壳成为长圆锥形,这种体形不利于动物在水中的运动及生存,于是逐渐地出现了由内脏囊的顶端开始做平面盘旋,后形成的外壳包围了先形成的外壳,这时壳与内脏囊仍对称。
前鳃类:壳的螺旋与倾斜虽解决了内脏囊隆起所带来的问题,但结果外套腔被压在了壳下,水流的通畅、鳃的呼吸作用及排泄物、生殖细胞等的排出都受到障碍,于是腹足类在进化中又出现了扭转现象,使身体扭转180°。这种扭转仅发生在外套膜及内脏囊部分,头、足不受影响,这种扭转或是顺时针方向或逆时针方向,扭转的结果使位于身体后端的外套腔移到了身体前端;原来平行连接的侧脏神经索也扭转形成8字形;左右侧的器官也交换了位置。
后鳃类:在进化中经过了扭转之后,又发生了反扭转,这就是后鳃类。反扭转的结果外套腔又回到了身体的后方,而且大部分种类外套腔消失,鳃也消失,而出现了次生性的皮肤鳃,贝壳也逐渐退化。侧脏神经索不再成8字形,但由于扭转而消失的一侧器官(鳃、心耳、肾)不再因反扭转而恢复。随着不同程度的反扭转,外套腔与壳也出现不同程度的退化,到裸鳃目时,外套腔及壳、本鳃都已消失,身体又变成了蠕虫状,外表又出现了两侧对称。
腹足类的壳为典型的螺旋圆锥形壳,壳顶是壳最先形成的部分,由壳顶围绕中心壳轴连续放大形成的螺形环称为螺层,最后形成的螺层为体螺层,其开口为壳口。除体螺层之外,其他螺层称螺旋部。
如果以壳顶向前,壳口面向观察者,壳口在壳轴的右侧,则称为右旋壳,壳口在壳轴的左侧,称左旋壳。大多数腹足类动物为右旋壳,少数种为左旋壳,也还有少数种同时具有两种个体。
大多数前鳃类足的后端背面,有一圆形角质板,或石灰质板,它的大小、形状与壳口完全一致,当头、足缩回壳内之后,这个板十分严密的完全封闭壳口,此壳板称为厣板,具有保护作用。
有的壳螺旋部不显著,成年的壳仅有体螺层极度膨大,如鲍
有的壳形有表现出两侧对称,如帽贝
有的壳面长出骨刺,如骨螺
有的壳完全埋在外套膜中,如泥螺、壳蛞蝓
有的壳完全消失,如海牛
大多腹足类是以足部肌肉的收缩来推动身体前进
一些小型生活在软质沙底的腹足类,可以靠足部纤毛运动推动身体前进,例如蜗牛、椎实螺等
在沙中穴居的腹足类,运动时靠足部充血形成犁或锚,然后拖动身体前进,如笋螺
水生后鳃类靠身体侧缘的波状收缩,或足部特化成翼在水中游泳运动,如琥螺
原始的腹足类,如原始腹足目,还具有一对双栉鳃,为了避免排出物对鳃及外套腔的污染,一些种类在外套膜及壳出现裂缝或小孔,水流经过鳃之后再将排出物带走
绝大多数的腹足类属于中腹足目,例如圆田螺、及新腹足目,例如骨螺,原始的双栉鳃已大部分消失,而代之以单栉鳃,仅鳃轴的一侧具有鳃丝
新腹足目外套膜前端延伸并卷拢形成水管,可以通过出水管将代谢物带走,这种鳃的结构、水管及水流方式适合于沙质水底表面生活或穴居,大量的海产前鳃类都是这种生活方式
在最原始的后鳃类,例如捻螺、泡螺仍具有圆锥形壳,仍有一单栉鳃,它的水流及呼吸方式仍相似于前鳃类
在较高等的后鳃类中,如海兔,其壳减小被包在外套膜中,外套腔及鳃仍然存在,但大大减小
裸鳃目中,如蓑海牛的壳、外套腔、鳃均已消失,而代之以身体的皮肤形成许多突起形成次生性皮肤鳃也称为裸鳃,以进行呼吸
肺螺类除了身体右侧有外套膜形成的一个气孔之外,外套腔已完全封闭,本鳃消失,由外套膜壁高度充血形成血管网,所谓“肺”
“肺”其实是靠外套膜底部的动以造成气体在外套膜腔中的流通,肺螺类即用这种方式进行气体交换,即使水生种类也是如此
当气候干燥或炎热的季节,蜗牛甚至躲在阴湿的环境下分泌粘液封闭壳口,以度过恶劣环境
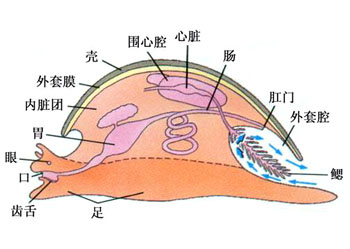
腹足类的消化系统由口、食道、胃、肠、肛门等器官组成;有的水生种类还出现了嗉囊及砂囊
植食性是腹足类中最重要的取食习性,植食性的腹足类是以各种海藻,水生或陆生植物为食,根据不同的生活环境而取食不同的植物
腹足类是以齿舌做为刮取食物的器官,齿舌的取食方式也因种不同,具有锉、刷、刀及运输器官等多种机能
法螺肉食性种类的齿舌上细齿的数目减少,但硬度增加,齿舌末端有倒勾、棘等,适合于切割及撕裂食物
芋螺的齿变成了单针状,中央具沟,末端具倒刺、基部有毒腺,其分泌物可以麻醉及杀死捕获物,甚至能使人致死
有一些肉食性腹足类具有钻孔取食的习性,如中玉螺、骨螺、红螺等,足端腺体可分泌酸性物质,以溶解壳的有机质及钙质成分
腐食性或沉积取食者,例如织纹螺、螺等,它们以潮汐带来的沉积物碎屑或腐烂的有机物颗粒为食
一些管居的蠕虫状的螺类,例如小蛇螺可以利用足腺分泌粘液形成粘液丝,分布在壳口周围,藉以诱捕浮游生物
光螺寄生于棘皮动物体壁上,其壳仍存在,足已高度退化,它的吻特别发达,以利于吸食寄主的体液及组织
前鳃类及肺螺类血浆中有呼吸色素即血蓝素,是一种含铜的蛋白质
海牛的内脏团
腹足类的头部明显,凤螺类具有很长的眼柄
后鳃类、肺螺类及极少数的前鳃类为雌雄同体,异体受精。交配时雄性生殖管末端膨大形成阴茎;雌管分出交配囊,粘腺及射囊
陆生腹足类交配时,常是互赠精囊到对方,交配后,精囊到对方的交配囊中释放出游离的精子,然后精子游到受精囊中使卵受精
水生腹足类产卵在水草或其他物体上,陆生种类产卵在潮湿土壤、洞穴等阴湿地方
蜗牛的卵
水生腹足类的卵
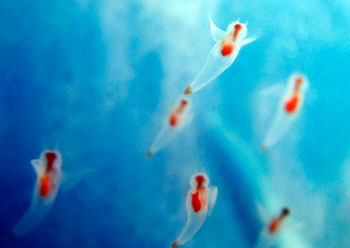
海产的腹足类绝大多数都有自由游泳的面盘幼虫期