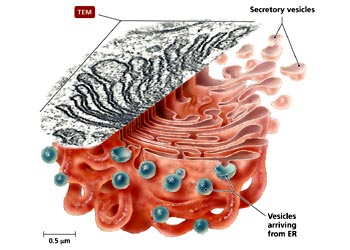
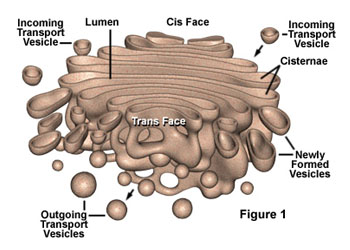
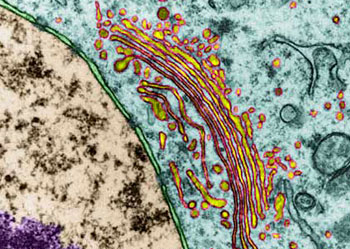
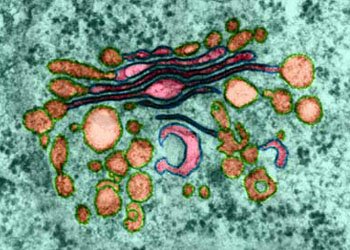
虽然早期的形态学观察结果就提示了高尔基体可能与细胞分泌活动有关,但对这一功能的了解却经历了一个较长的逐渐认识的过程。70年代初,Caro用3H-亮氨酸对胰腺的腺泡细胞进行脉冲标记,发现在脉冲标记3分钟后,放射自显影银粒主要位于内质网; 20分钟后,银粒出现在高尔基体; 120分钟后则位于分泌泡并开始在顶端释放。实验显示了分泌性蛋白在细胞内的合成与转运途径,其转运的过程是通过高尔基体来完成的,后来的研究进一步表明,除分泌性蛋白外,很多细胞质膜上的膜蛋白、溶酶体中的酸性水解酶及胶原纤维等胞外基质成分都是通过高尔基体完成其定向转运过程。作为蛋白质合成主要场所的内质网常常同时合成多种蛋白质,那么高尔基体怎样完成对这些蛋白质的分类与转运功能呢?60年代,人们发现溶酶体中所有的酶都有共同的标志。70年代证明这一共同标志就是6-磷酸甘露糖(M6P),80年代纯化了与这一反应有关的酶及M6P受体,从而把溶酶体酶在高尔基体中的分类过程作为了解高尔基体功能的一个重要例子。溶酶体中含有几十种酸性水解酶类,它们在内质网上合成后进入高尔基体。在内质网上合成时发生了N―连接的糖基化修饰,即把一个寡糖链共价结合到溶酶体酶分子中的天冬酰氨残基上。在高尔基体的顺面的膜囊中存在N-乙酰葡萄糖胺磷酸转移酶和N-乙酰葡萄糖胺磷酸糖苷酶,在这二种酶的催化作用下,寡糖链中的甘露糖残基磷酸化产生6-磷酸甘露糖。这种特异的反应,只发生在溶酶体的酶上,而不发生在其它的糖蛋白上,估计溶酶体酶本身的构象含有某种磷酸化的信号,如改变其构象则不能被识别也就不能形成6-磷酸甘露糖。在高尔基体反面的膜囊上结合着6-磷酸甘露糖的受体,由于溶酶体酶的许多位点上都可形成6-磷酸甘露糖,从而大大增加了与受体的亲和力,这种特异的亲和力使溶酶体的酶与其它蛋白质分离并起到局部浓缩的作用。在一种称为I细胞(inclusion cell)病中,病人由于N-乙酰葡萄糖胺磷酸转移酶单基因的缺损,因此不能合成6-磷酸甘露糖,溶酶体的酶也就不能被受体识别,因而无法转运到溶酶体中。在内质网合成的蛋白质很多都是糖蛋白,而且这些蛋白质的糖链在高尔基体中经历十分复杂的修饰。于是人们猜测这种修饰作用可能与蛋白质在高尔基体中的分类有关。然而用DNA重组技术证明,多种糖蛋白在去掉糖链后仍能正常地输送到细胞的特定部位,说明糖链在多数蛋白质的分类中并不起决定性的作用。上述溶酶体酶的分选途径可能仅仅是一个特例,况且也不是溶酶体酶唯一的途径,已发现在肝细胞中溶酶体酶还存在不依赖于6-磷酸甘露糖的另一种分选途径。一个很有趣的实验显示了蛋白质在高尔基体中分选及其转运的信息仅存在于编码这个蛋白质的基因本身。流感病毒和水泡性口炎病毒可同时感染上皮细胞,这两种有囊膜病毒的囊膜蛋白均在内质网上合成,然后经高尔基体转运到细胞质膜上。流感病毒的囊膜蛋白特异性地转运到上皮细胞游离端的细胞质膜上,而水泡性口炎病毒的囊膜蛋白则转运到基底面的细胞质膜上。将克隆的流感病毒囊膜蛋白的基因和水泡性口炎病毒囊膜蛋白的基因同时在上皮细胞中表达,结果显示,两种病毒囊膜蛋白的合成、转运途径及在细胞质膜上的分布与两种病毒同时感染细胞时,病毒囊膜蛋白在质膜上的分布相同。目前,人们发现水泡性口炎病毒囊膜蛋白在由内质网合成后进入高尔基体时,存在于细胞质基质一侧的双酸分选信号(Asp-x-Gln或DxE)起重要的作用,其它一些膜蛋白也具有这一信号序列,表明膜蛋白在由内质网向高尔基体转运时,也存在一种选择性的转运机制。但是关于高尔基体对各种蛋白自身所携带的分选信号的识别、进而对其分类、包装与运送的机制,目前还不很清楚。至于高尔基体对蛋白转运的调控机制目前了解甚少。然而,一些实验显示一类小分子的G蛋白(GTP binding regulatory protein or G protein)Rab在高尔基体的囊泡转运中起重要调节作用。