细胞膜细胞核 线粒体叶绿体内质网 核糖体高尔基体 溶酶体微体微丝微管 中间纤维
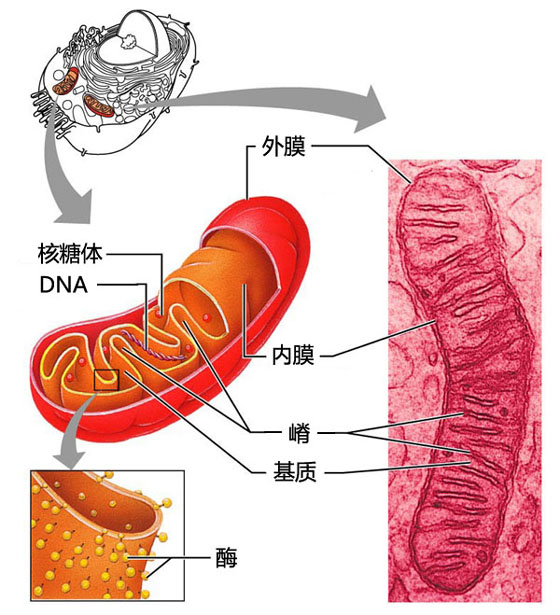
线粒体
线粒体的形态、大小、数量与分布,在不同细胞内变动很大,就是同一细胞在不同生理状态下也不一样。线粒体在生活细胞中具有多形性、易变性、运动性和适应性等特点。其形状多种多样,但以线状和颗粒状最常见,也可呈环形、哑铃形、分枝状或其它形状。 在一定条件下线粒体的形状变化是可逆的,线粒体一般直径为0.5~1.0μm,长1.5~3.0μm。其大小也因细胞种类和生理状况的不同而不一样。例如,大鼠肝细胞的线粒体可长达5μm。在胰腺的外分泌细胞中可观察到巨大线粒体,其长达10~20μm。人的成纤维细胞线粒体甚至可长达40μm。线粒体的形状与大小并不是固定的,而是随着代谢条件的不同而改变,它可能反映线粒体处于不同的代谢状态。一般动物细胞内线粒体的数目由数百到数千个,肝细胞内约有1 700个左右。植物细胞的线粒体数量一般较动物细胞的少。线粒体的数目在不同类型细胞内有很大差别,许多哺乳动物成熟的红细胞缺少线粒体,利什曼原虫中只有一个巨大的线粒体,海胆卵母细胞则多达30万个,巨大变形虫可高达50万个。线粒体的数目还与细胞的生理功能及生理状态有关,在新陈代谢旺盛的细胞中线粒体多,如人和哺乳动物的心肌、小肠、肝等内脏细胞中线粒体很丰富;飞翔鸟类胸肌细胞线粒体的数目比不飞翔鸟类的多;运动员的肌细胞线粒体比不常运动的人多。线粒体在细胞内的分布一般是不均匀的,例如,肠上皮细胞中的线粒体呈两极性分布,集中于顶部和基部。根据细胞代谢的需要,线粒体可在细胞质中运动、变形和分裂增殖,如在玉米的小孢子发育过程中,线粒体定向的运动、聚集与分散,在此过程中绒毡层细胞中线粒体的数量可增加40多倍。线粒体往往在细胞代谢旺盛的需能部位比较集中,如分泌细胞的线粒体聚集在分泌物合成的区域;肌细胞的线粒体沿肌原纤维规则排列;精子细胞的线粒体集中在鞭毛中区。线粒体的这种分布显然有利于需能部位的能量供应。线粒体在细胞质中迁移时,往往与微管有关。
线粒体的化学组成及酶的定位