细胞膜细胞核 线粒体叶绿体内质网 核糖体高尔基体 溶酶体微体微丝微管 中间纤维
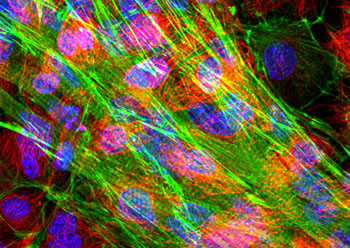
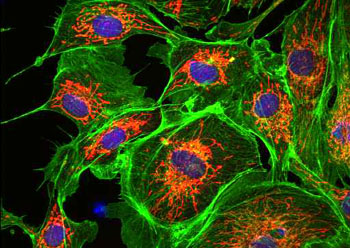
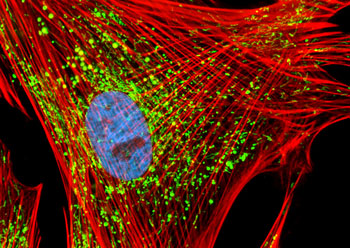
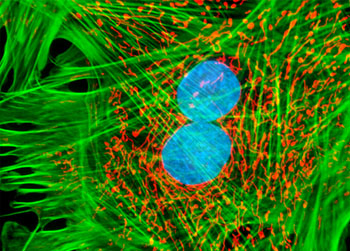
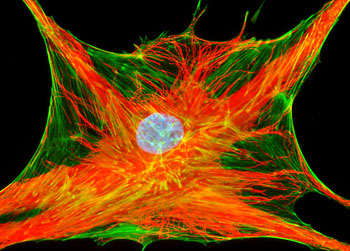
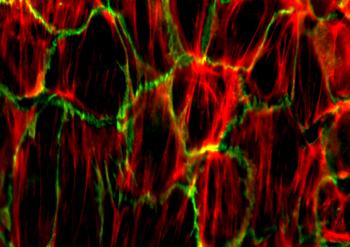
微丝
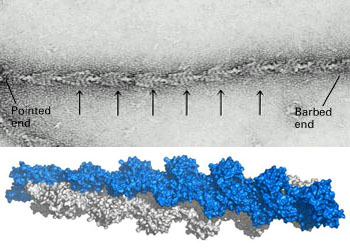
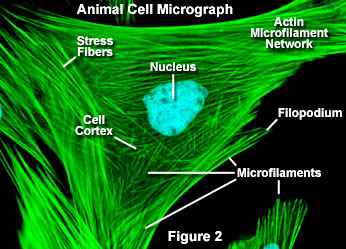
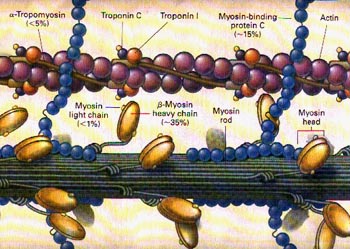
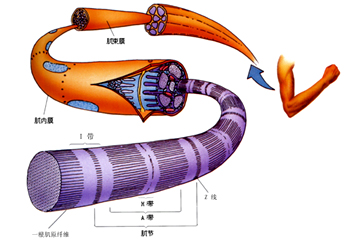
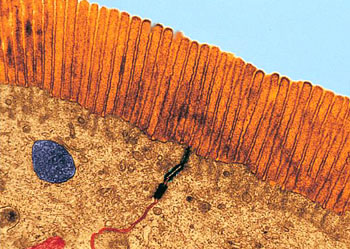
微丝,亦称为纤维形肌动蛋白(F-actin),是由球形肌动蛋白(G-actin)单体形成的多聚体。肌动蛋白单体具有极性,装配时呈头尾相接,故微丝也具有极性。比较传统的模型认为微丝是由两条肌动蛋白单链呈右手螺旋盘绕形成的纤维,近年来则认为微丝是由一条肌动蛋白单体链形成的右手螺旋,每个肌动蛋白单体周围都有4个单体,呈上、下及两侧排列。在含有ATP和Ca2+以及很低浓度的Na+、K+等阳离子溶液中,微丝趋于解聚成G-actin;而在Mg2+和高浓度的Na+,K+溶液诱导下,G-actin则装配为纤维状肌动蛋白,新的G-actin加到微丝末端,使微丝延伸。G-actin可以加到微丝两端,但(+)极组装的速度较(-)极快,在一定条件下,微丝可以表现出一端因加亚单位而延长,而另一端因亚单位脱落而减短,这种现象称为踏车行为(tread milling)。微丝在结构上的极性对行使其功能也是必要的。肌动蛋白可在体外装配成微丝,其结构与细胞中分离的微丝相同,可以通过聚合-解聚纯化微丝。在体内,有些微丝是永久性的结构,如肌肉中的细丝及肠上皮细胞微绒毛中的轴心微丝等;有些微丝是暂时性的结构,如胞质分裂环中的微丝。血小板激活及无脊椎动物精子细胞顶体反应中的微丝只有在需要时方进行装配。实际上,在大多数非肌肉细胞中,微丝是一种动态结构,持续进行组装和解聚,并与细胞形态维持及细胞运动有关。体内肌动蛋白的装配在两个水平上受到结合蛋白的调节:(1)游离肌动蛋白单体的浓度;(2)微丝横向连接成束或成网的程度。细胞内许多微丝结合蛋白参与调节肌动蛋白的组装。