细胞膜细胞核 线粒体叶绿体内质网 核糖体高尔基体 溶酶体微体微丝微管 中间纤维
溶酶体
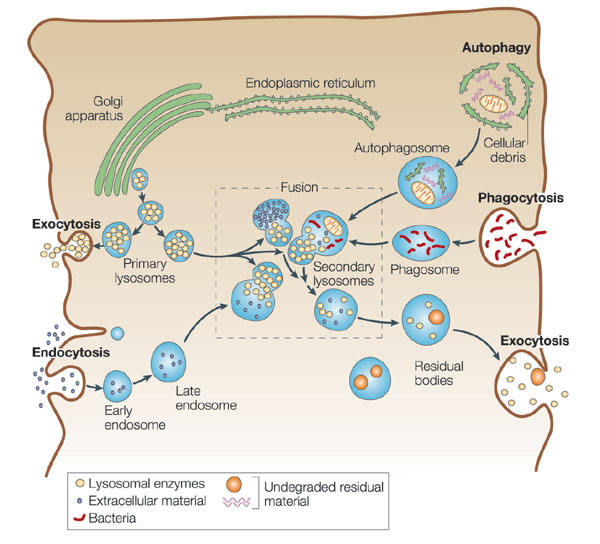
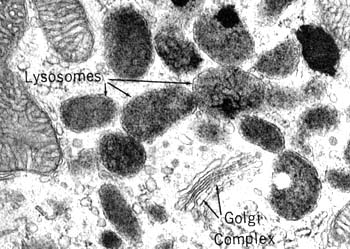
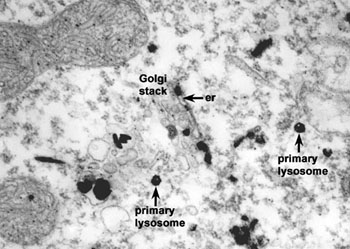
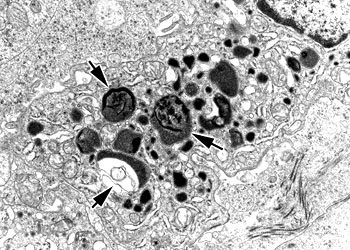
溶酶体是一种异质性(heterogenous)的细胞器,这是指不同的溶酶体的形态大小,甚至其中所包含的水解酶的种类都可能有很大的不同,根据溶酶体处于完成其生理功能的不同阶段,大致可分为初级溶酶体(primary lysosome)、次级溶酶体(secondary lysosome) 和残余小体(residual body)。初级溶酶体呈球形,直径约0.2~0.5μm,内容物均一,不含有明显的颗粒物质,外面由一层脂蛋白膜围绕,厚度为7.5nm。其中含有多种水解酶类,如蛋白酶、核酸酶、糖苷酶、脂酶、磷脂酶、磷酸酶和硫酸酶等,其共同的特征是都属酸性水解酶,即酶的最适pH为5左右。如将氢氧化氨或氯奎(chloroquine)等可穿入细胞膜的碱性物质加入细胞培养液中,致溶酶体中pH值提高至7左右,则可使溶酶体酶失去活性。溶酶体膜在成分上也与其它生物膜不同。(1)嵌有质子泵,借助水解ATP释放出的能量将H+泵入溶酶体内,使溶酶体中的H+ 浓度比细胞质中高100倍以上,以形成和维持酸性的内环境;(2)具有多种载体蛋白用于水解的产物向外转运;(3)膜蛋白高度糖基化,可能有利于防止自身膜蛋白的降解。目前已发现60余种溶酶体的酶类,多数为可溶性的酶,有些整合在溶酶体膜上。酶蛋白本身的结构能抗御酸变性作用。已克隆了近20种酶的cDNA,并测出了一些酶的基因序列,发现溶酶体的酶具有某些特征的同源序列。此外,催化相关反应的某种溶酶体的酶和非溶酶体酶之间蛋白质一级结构也非常相似,甚至与低等真核生物及原核生物的有关酶也非常相似。显然,溶酶体的酶与相关的非溶酶体酶是属于结构与功能上相似的一类酶的家族,推测它们在进化中有共同的起源。次级溶酶体是初级溶酶体与细胞内的自噬泡或异噬泡、胞饮泡或吞噬泡融合形成的复合体,分别称之为自噬溶酶体(autophagolysosome)和异噬溶酶体(phagolysosome),二者都是进行消化作用的溶酶体。次级溶酶体中可能包含多种生物大分子、颗粒性物质、线粒体等细胞器乃至细菌等,因此其形态不规则,直径可达几个微米。电镜显示其内部结构非常复杂,常含有颗粒、膜片甚至某些细胞器。经过一段时间的消化后,小分子物质可通过膜上的载体蛋白转运到细胞质基质中,供细胞代谢使用,未被消化的物质残存在溶酶体中形成残余小体或称后溶酶体。残余小体可通过类似胞吐的方式将内容物排出细胞。用溶酶体的标志酶反应,可辨认出不同形态与大小的溶酶体。酸性磷酸酶(acid phosphatase)是常用的标志酶,用这种方法不仅有助于研究溶酶体的发生与成熟过程,而且还发现了多泡体、线状溶酶体等多种类型的溶酶体,但其机能尚不完全清楚。因此溶酶体可以看作是以含有大量酸性水解酶为共同特征的,不同形态大小,执行不同生理功能的一类异质性的细胞器。少量的溶酶体酶泄露到细胞质基质中,并不会引起细胞损伤,其主要原因是细胞质基质中的pH值为7.0左右,在这种环境中溶酶体酶的活性大大降低。此外,在酵母细胞质中已发现一些蛋白可以特异地与溶酶体酶结合而使其丧失活性。植物细胞的液泡中含有多种水解酶类,具有与动物细胞溶酶体类似的功能,一般液泡约占细胞总体积的30%以上,但在不同细胞中液泡体积从5%直至90%不等。除此之外,液泡还具有储存营养与废物、调节细胞体积增长及细胞膨压等多种作用。


.jpg)