细胞膜细胞核 线粒体叶绿体内质网 核糖体高尔基体 溶酶体微体微丝微管 中间纤维
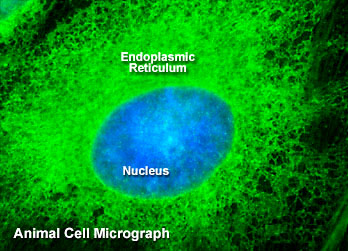
内质网
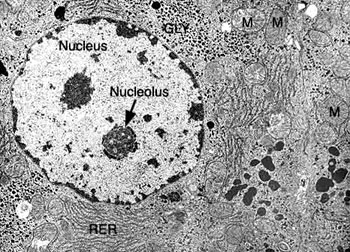
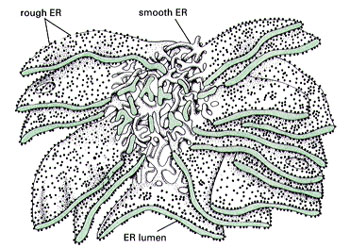
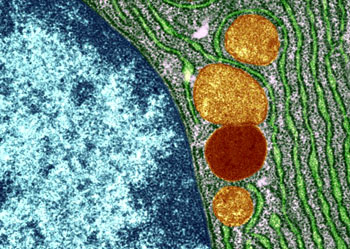
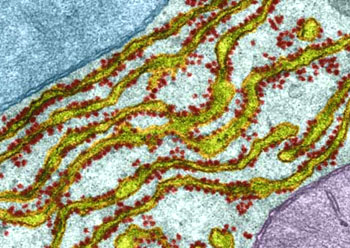
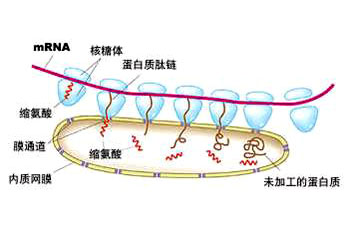
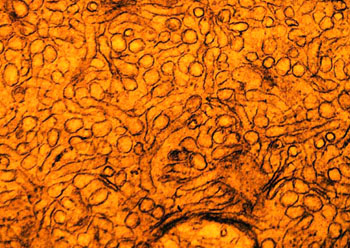
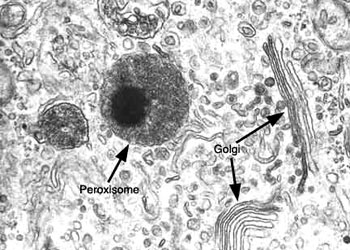
根据结构与功能,内质网可分为两种基本类型: 粗面内质网( rough endoplasmic reticulum,rER)和光面内质网(smooth endoplasmic reticulum,sER)。粗面内质网多呈扁囊状,排列较为整齐,因在其膜表面分布着大量的核糖体而命名。它是内质网与核糖体共同形成的复合机能结构,其主要功能是合成分泌性的蛋白和多种膜蛋白。因此在分泌细胞(如胰腺腺泡细胞)和分泌抗体的浆细胞中,粗面内质网非常发达),而在一些未分化的细胞与肿瘤细胞中则较为稀少。内质网膜上有一种称为移位子(translocon)的蛋白复合体,直径约8.5nm,中心有一个直径为2nm的“通道”,其功能与新合成的多肽进入内质网有关,在哺乳动物细胞中,移位子的主要成分是与蛋白分泌相关的一种多肽Sec61p等组成复合物。表面没有核糖体结合的内质网称光面内质网,光面内质网常为分支管状,形成较为复杂的立体结构。光面内质网是脂类合成的重要场所,细胞中几乎不含有纯的光面内质网,它们只是作为内质网这一连续结构的一部分。光面内质网所占的区域通常较小,往往作为出芽的位点,将内质网上合成的蛋白质或脂类转移到高尔基体内。在某些细胞中,光面内质网非常发达并具有特殊的功能,如合成固醇类激素的细胞及肝细胞等。用密度梯度离心技术可将肝细胞中的光面内质网和粗面内质网分离出来,发现粗面内质网上有20余种与光面内质网上不同的蛋白质。既然内质网是一个连续的整体结构,因此,在内质网膜上可能有某些特殊的装置,将光面内质网与粗面内质网的部位间隔开来,并维持其形态。否则在内质网膜这个二维的流体结构中,不同区域的脂类和蛋白质就会因侧向扩散而趋于平衡。内质网与其它细胞器关系的研究,对阐明细胞的一些生理生化过程及细胞器的发生与进化是很有意义的,也提出一些有启示性的设想。超微结构研究表明,向内折叠的细胞质膜有时与内质网相连接,甚至有管道相通。原核细胞的细胞质膜内侧有时附着大量核糖体,因而一些人认为在细胞进化过程中,内质网可能由细胞质膜演化而来。内质网膜常与外层核膜连接,内质网的腔与核周隙相沟通,而且外核膜有时也附着大量的核糖体,这种结构上的联系不仅反映了核-质间的物质交换,同时也提出了内质网与核膜在发生上的同源关系。光面内质网与高尔基体在结构、功能与发生上的关系更为密切。此外,在合成旺盛的细胞内,粗面内质网总是与线粒体紧密相依,过去的解释认为线粒体是内质网执行功能时所需能量的直接“供应站”。最近发现脂类的相互交换及Ca2+释放的调节都与之密切相关。在间期细胞中,内质网的分布常常与微管的走向一致,且总是沿微管向细胞周缘延伸。已发现一种微管马达蛋白――驱动蛋白(kinesin)与内质网结合,推测既然内质网一端固定在核膜上,另一端在驱动蛋白的牵引下沿微管向外延伸形成复杂的网状结构。内质网对外界因素的作用(如射线、化学毒物、病毒等)非常敏感,粗面内质网发生的最普遍的病理变化是内质网腔扩大并形成空泡,继而核糖体从内质网膜上脱落,蛋白质合成受阻。