细胞膜
细胞核
线粒体
叶绿体
内质网
核糖体
高尔基体
溶酶体
微体
微丝
微管
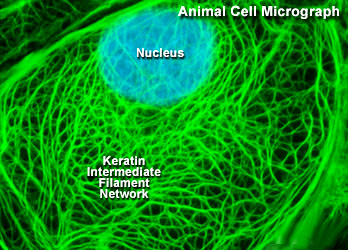
中间纤维
你的位置:
首页
>
细胞
> 中间纤维
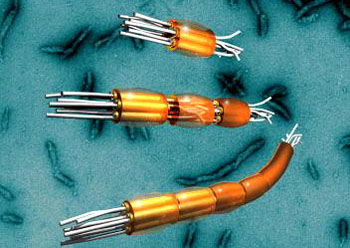
60年代中期,在哺乳动物细胞中发现10nm纤维,因其直径介于肌粗丝和细丝之间,故被命名为中间纤维(intermediate filament,IF),又称中间丝。中间纤维的成分比微丝和微管复杂,按其组织来源及免疫原性可分为5类:(1)角蛋白纤维(keratin filament),存在于上皮细胞中;(2)波形纤维(vimentin filament),存在于间质细胞和中胚层来源的细胞中;(3)结蛋白纤维(desmin filament),存在于肌细胞中;(4)神经元纤维(neurofilament),存在于神经元中;(5)神经胶质纤维(neuroglial filament),存在于神经胶质细胞。中间纤维的分布具有严格的组织特异性。这一点已被应用于肿瘤临床鉴别诊断,以鉴别肿瘤细胞的组织来源,显示了良好的应用前景。中间纤维蛋白的氨基酸顺序已基本清楚,许多肽链序列已经测定。研究发现中间纤维蛋白来源于同一基因家族,具有高度同源性。中间纤维蛋白结构特征如图9-20所示,在中间纤维蛋白分子肽链中部有一段约310个氨基酸残基的α-螺旋区是高度保守的。两个相邻亚基对应的α-螺旋区形成双股超螺旋,即40~50nm的杆部(rod),组装为中间纤维的主干(backbone),这是5类中间纤维的共同结构特征。同一型的中间纤维蛋白其杆状区氨基酸顺序有70~90%的同源性,但不同型的中间纤维蛋白的同源性则低于30%。中间纤维蛋白杆状区由螺旋1和螺旋2构成,螺旋1和螺旋2的长度约为22nm,杆状区长度约在47nm左右。螺旋1和螺旋2又可分为A、B两个亚区,4个螺旋区间由3个短片段相连接,其中L12连接螺旋1和螺旋2,L1和L2分别连接1A与1B和2A与2B。而中间纤维蛋白分子非螺旋化的头部(N端)和尾部(C端)的氨基酸顺序和肽链长度在各类不同中间纤维蛋白分子中有很大差异。对于每一特定类型的中间纤维蛋白,其头部和尾部又可进一步分为不同的亚区,(1)H亚区:同源区;(2)V亚区:可变区;(3)E亚区:末端区。根据中间纤维蛋白的氨基酸顺序的同源性,近年对中间纤维成分又提出新的分类:Ⅰ.酸性角蛋白;Ⅱ.中性和碱性角蛋白; Ⅲ.波形纤维蛋白,结蛋白,胶质纤维酸性蛋白;57kD神经中间纤维蛋白;Ⅳ.神经元纤维蛋白;Ⅴ.核纤层蛋白(核内中间纤维蛋白,见后文);Ⅵ.nestin,一种在神经干细胞中发现的蛋白,根据其一级结构确定为中间纤维蛋白。
中间纤维
IF
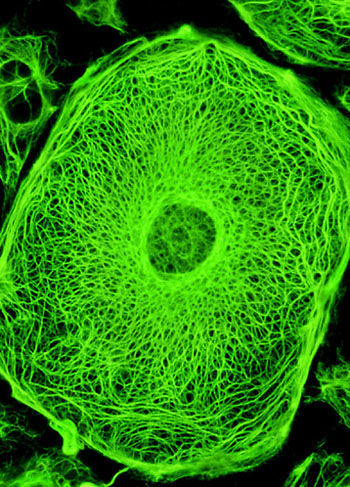
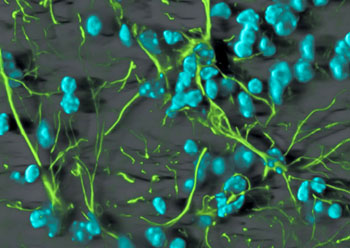
鼠袋鼠细胞中间纤维电镜照
神经元中间纤维电镜照
星形胶质细胞中间纤维电镜照
中间纤维蛋白基因
目前对中间纤维蛋白基因已有所了解,中间纤维蛋白基因含有内含子/外显子结构,同型中间纤维蛋白基因具有几乎一致的内含子/外显子结构,Ⅰ-Ⅲ型中间纤维蛋白基因在其顺序组织上相似,Ⅳ型中间纤维蛋白基因在其顺序组织上与Ⅰ-Ⅲ型中间纤维基因有较大差别。中间纤维蛋白基因定位和调节序列的性质尚不清楚。中间纤维蛋白基因表达:蛋白表达方式已经清楚,中间纤维蛋白的表达具有严格的组织特异性。中间纤维基因蛋白表达的组织和发育调节可能是在转录和转录后水平,但其规律尚不清楚。
中间纤维的装配
中间纤维蛋白的杆部组装为中间纤维的主干(backbone)。首先,两个相邻亚基的对应α-螺旋区形成双股超螺旋,即二聚体。研究表明,二聚体中中间纤维蛋白平行排列,50nm长。Ⅰ-Ⅱ型中间纤维蛋白分子(角蛋白)组装为异二聚体(het-erodimer),尽管在体外组装中同二聚体是可能的。一般由一种ⅠⅡ型角蛋白和一种与之相配对的Ⅱ型角蛋白组装成二聚体。Ⅲ-Ⅳ型中间纤维蛋白分子由相同的分子组装为二聚体,即同二聚体(isodimer)。中间纤维组装的第二步是二聚体怎样组装为四聚体(tetramer),早期曾提出中间纤维组装的三股螺旋模型,现已排除了这种可能。目前认为由两对超螺旋形成四聚体,四聚体可能是中间纤维解聚的最小亚单位。对四聚体中分子如何排列尚有争议,可能是平头(50nm)或半交叠(70nn),很可能是反向平行排列(例如,中间纤维没有极性),但也不排除顺向平行排列的可能。关于反向平行和半分子交叠发生在四聚体内还是四聚体间,尚有待进一步研究。由四聚体或八聚体进一步组装为中间纤维的确切方式尚不清楚,组装好的中间纤维具有多态性,最多见的是由8个四聚体或4个8聚体组装为中间纤维,中间纤维横截面可由32个分子组成;也有一些中间纤维,在横截面上包含5~6个或11~12个四聚体。此外,中间纤维可能有原纤维(4聚体,protofilament)或原丝(8聚体,protofibril)等中间类型。中间纤维蛋白杆部形成中间纤维的核心,直径为8~9nm。中间纤维蛋白的末端大多凸出于中间纤维的核心之外,中间纤维22nm或48nm的纵向周期与末端区域突出有关。中间纤维真正的直径取决于肽链末端的大小,通常为12~16nm,由于外侧2~3nm的密度较低,电镜下难以分辨,故中间纤维在电镜下一般为10nm左右。由中间纤维核心伸出的末端区域可能和中间纤维与细胞内结构相互作用及功能有关。
中间纤维是中空的管状结构, 直径10nm,介于肌粗丝和细丝之间,故名。其功能并不十分清楚。一般认为,中间纤维在细胞质、细胞间和组织中起支架作用,并与细胞核定位有关系
在电镜下,中间纤维的电子密度很低。中间纤维具有严格的组织特异性,在不同组织细胞中,化学组成不同,有角蛋白纤维、波形纤维、结蛋白纤维、神经元纤维、神经胶质纤维等5种类型
一般认为组装中间纤维时,先是通过2条多肽链杆部的α-螺旋区结合形成二聚体,1对二聚体聚合为四聚体,2个四聚体再形成一根亚丝,由4根亚丝盘绕成1根完整的中间纤维
中间纤维结合蛋白
中间纤维结合蛋白(intermediate filament associated protein,IFAP)是一类在结构和功能上与中间纤维有密切联系,但其本身不是中间纤维结构组分的蛋白。IFAP可能在中间纤维上或其两端与中间纤维紧密或松散结合。可能作为细胞中中间纤维超分子结构的调节者。确定IFAP的标准:(1)在细胞内与中间纤维共分布,(2)抗高盐与非离子去垢剂抽提,与中间纤维共同分离,(3)与中间纤维经历相同的解聚和重组装周期,(4)在体外能与由纯中间纤维蛋白亚基组装的中间纤维结合。迄今为止已报道了大约15种IFAP:filaggrin、IFAP(~300kD)、IFAP(70~280kD)、plectin(300kD)、synemin(230kD)、p95、paranemin(280kD)、epinemin(44.5kD)、p50(50kD)、NAPA73(73kD)等。其它一些蛋白也具有IFAP的性质。如桥板蛋白(desmoplakin)1和2参与桥粒形成;微管结合蛋白(MAP2)参与中间纤维与微管间横桥;血影蛋白参与中间纤维与膜结合;锚蛋白参与中间纤维与膜结合。已了解的IFAP的一些特性可归结如下:(1)已经分离的IFAP具有中间纤维类型特异性;(2)IFAP的表达有细胞专一性;(3)不同的IFAP可存在于同一个细胞中,与不同的中间纤维组织状态相联系;(4)在细胞中某些IFAP的表达与细胞的功能和发育状态有关。filaggrin的功能是使角蛋白纤维聚集,在角质化细胞中形成大的纤维聚集物,该蛋白的表达是角质化的分化特异性标志。除filaggrin以外,大多中间纤维结合蛋白的功能尚不清楚。
中间纤维的组织及动态变化
与微丝、微管不同,中间纤维蛋白合成后,基本上均组装为中间纤维,游离的单体很少。细胞内的中间纤维蛋白均受到不同程度的化学修饰,包括乙酰化,磷酸化等,可能与中间纤维的动态变化及功能活动有关。对细胞中中间纤维的组织尚了解不多在同一个细胞中,中间纤维有不同的组织结构,如散在的纤维,复杂的网络结构,粗大的中间纤维束,密集的中间纤维“邻核帽”(juxta-nuclearcap)等,中间纤维在胞质中形成精细发达的纤维网络,外与细胞膜及细胞外基质相连,内与核纤层有直接联系,目前对中间纤维与核膜和细胞表面联系的认识仍是初步的。在胞质中中间纤维与微丝、微管及其它细胞器有错综复杂的纤维联络。中间纤维与其它骨架组分确切关系尚知之不多。尽管中间纤维在细胞中是相当稳定的结构,在一定的生理条件或实验条件下,中间纤维或中间纤维网会发生改变,如有丝分裂过程中中间纤维发生解聚和重组装。其它如胰酶消化、秋水仙素处理、显微注射中间纤维蛋白抗体、热休克、病毒感染、酒精肝、Alzhemer’s病都可对细胞内中间纤维网的组织状态产生影响,使中间纤维网崩塌。迄今对组织中的中间纤维的结构组织仅有一些概念性的了解,中间纤维通过桥粒在细胞间连续,推测中间纤维跨细胞的结构,对维持上皮的连续性是重要的。
中间纤维与细胞分化
微丝和微管在各种细胞中均相同,而中间纤维蛋白的表达具有组织特异性,中间纤维与细胞分化的关系非常密切,引起广泛重视。研究主要集中在胚胎发育和上皮分化两个方面。(1)胚胎发育:小鼠胚胎发育过程中,最初胚胎细胞中表达角蛋白,胚胎发育第8~9天,将要发育为间叶组织的细胞中,角蛋白表达下降并停止,同时出现波形纤维蛋白的表达。类似的表型变化见于神经外胚层发育中,首先是出现角蛋白的表达,第11天左右,角蛋白表达停止,波形纤维蛋白出现;一些将要发育为星形细胞的细胞在第18天左右开始同时表达波形纤维蛋白和胶质酸性蛋白,而另一些将要发育为神经细胞的细胞在表达角蛋白向波形纤维蛋白表达转变后短时间内即开始表达神经元纤维蛋白,继而波形纤维蛋白表达下降并停止。由此可见,胚胎细胞能根据其发育的方向调节中间纤维蛋白基因的表达。(2)上皮分化:上皮组织的分化过程中角蛋白表达的变化为研究中间纤维与细胞分化的关系提供了一个重要例证(图9-24),在上皮细胞中,酸性角蛋白(Ⅰ型)和中性-碱性角蛋白(Ⅱ型)成对表达,并具有特异性。如单层上皮细胞中特异表达K18(45kD)-K8(52kD)和K17(46kD)-K7(54kD)两对角蛋白;复层上皮细胞中特异表达K16(48kD)-K6(56kD)和K14(50kD)-K5(58kD)两对角蛋白;食管上皮细胞中特异表达K13(51kD)-K4(59kD)角蛋白对;角膜上皮细胞中特异表达K12(55kD)-K3(64kD)角蛋白对;皮肤细胞中特异表达K10(56.5kD)-K1-2(65~67kD)角蛋白对。角蛋白对中酸性角蛋白(Ⅰ型)和中性-碱性角蛋白(Ⅱ型)分子量通常相差8kD。迄今关于中间纤维在细胞分化中的变化已积累了很多资料,但有关中间纤维在细胞分化中的作用尚不清楚。
中间纤维在生物进化中的起源
自发现中间纤维以来,一般认为中间纤维在生物进化中的起源比微丝和微管晚,免疫细胞化学、电镜及生化研究显示中间纤维似乎局限于脊椎动物细胞,其进化上的保守性比微丝和微管要差。然而,近年来有一些报道显示在无脊椎动物细胞中可能也存在类中间纤维蛋白或10nm样纤维。植物细胞中是否存在中间纤维?尚是一个有待解决的问题,翟中和等以胶体金免疫电镜技术、免疫印迹技术及分子杂交技术对植物细胞类中间纤维结构,蛋白成分及其基因同源序列进行了初步研究,从多方面提供了植物细胞中存在类中间纤维的证据。从中间纤维蛋白一级结构及其基因的分析认为中间纤维蛋白应属于同一基因家族,由同一个祖先基因进化而来,并提出中间纤维蛋白基因进化模式(图9-25)。Ⅰ~Ⅲ型中间纤维蛋白与Ⅳ中间纤维蛋白较早分叉,从蛋白一级结构及其基因结构看,核纤层蛋白可能代表中间纤维蛋白较原始的形式。最近对无脊椎动物细胞中间纤维蛋白一级结构的研究表明,核纤层蛋白与无脊椎动物细胞中间纤维蛋白更为相似。
中间纤维的功能
中间纤维的功能至今仍是很不清楚的,一个重要的原因是迄今尚未找到一种中间纤维特异性工具药。一般认为,中间纤维在细胞质中起支架作用,并与细胞核定位有关。同时,中间纤维在细胞间或者组织中起支架作用,如角蛋白中间纤维参与了桥粒的形成和维持,结蛋白纤维参与了肌肉Z盘的构造,但上述支架功能都是基于形态学观察推测的。中间纤维在胞质中形成精细发达的纤维网络,外与细胞膜及细胞外基质相连,内与核纤层有直接联系,推测中间纤维参与传递细胞内机械的或分子的信息,可以是从细胞膜传至细胞核,也可以是由细胞核传至细胞膜。Traub(1985)提出中间纤维蛋白核功能的模型,其根据是波形纤维蛋白与DNA及组蛋白均有较强的亲和力,并发现Ca2+激活的中性硫醇蛋白酶和另一种Ca2+激活的水解酶可以优先水解波形纤维蛋白氨基端,使其失去装配成纤维的能力。中间纤维蛋白核功能假说的主要观点是:中间纤维网架直接伸到细胞膜,当外界信号如激素、外源凝集素、免疫球蛋白、生长因子等与细胞膜上受体作用时,即启动了Ca2+的流入,Ca2+浓度升高引起级联反应,导致Ca2+调酶的激活,中间纤维蛋白的N端被水解,其大分子产物失去装配成纤维的能力并进入细胞核内,通过与组蛋白和DNA的作用来调节复制和转录。近年来发现中间纤维与mRNA的运输有关,胞质mRNA锚定于中间纤维可能对其在细胞内的定位及是否翻译起决定作用。此外,神经元纤维在神经细胞轴突运输中起作用。