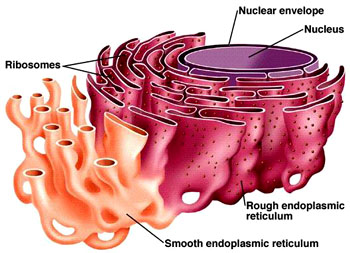
细胞膜细胞核 线粒体叶绿体内质网 核糖体高尔基体 溶酶体微体微丝微管 中间纤维
核糖体
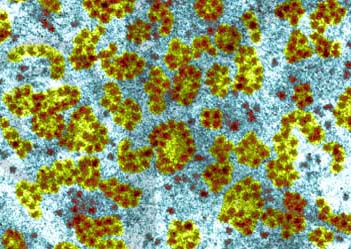
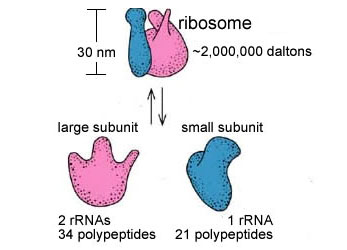
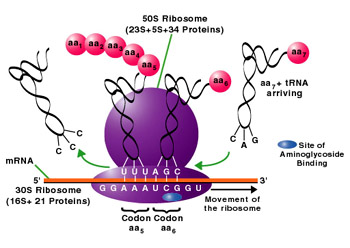
生物有机体细胞内有两种基本类型的核糖体:一种是70S(S为Sverdberg沉降系数单位)的核糖体,其相对分子量为2.5×106,原核细胞的核糖体为70S,真核细胞线粒体与叶绿体内的核糖体也近似于70S;另一种是80S的核糖体,相对分子量为4.8×106,真核细胞的核糖体(除线粒体与叶绿体核糖体外)均为80S。不论70S或80S的核糖体,均由大小不同的两个亚单位(subunit)构成。体外实验表明70S的核糖体在Mg2+浓度小于1m mol/L的溶液中,易离解为50S与30S的大小亚单位,当溶液中Mg2+浓度大于10m mol/L时,两个核糖体常常形成100S的二聚体。核糖体大小亚单位在细胞内常常游离于细胞质基质中,只有当小亚单位与mRNA结合后大亚单位才与小亚单位结合形成完整的核糖体。肽链合成终止后,大小亚单位解离,又游离存在于细胞质基质中。
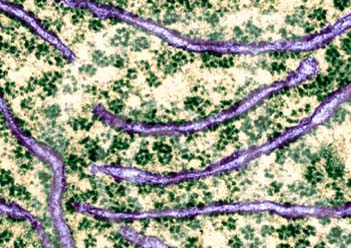
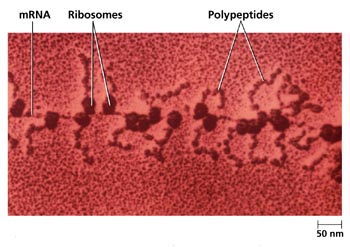
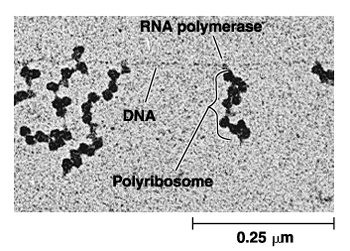
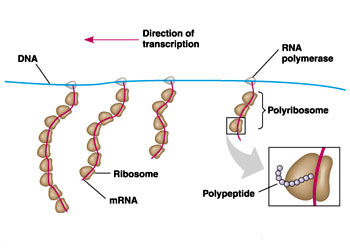
核糖体是蛋白质合成的机器。但核糖体在细胞内并不是单个独立地执行功能,而是由多个甚至几十个核糖体串连在一条mRNA分子上高效地进行肽链的合成。这种具有特殊功能与形态结构的核糖体与mRNA的聚合体称为多聚核糖体(polyribosome或polysome)。在细胞的超薄切片中, 不论是附着核糖体还是游离核糖体,经常可以看到它们串联排列成簇状、环状、串珠状,甚至雪花状等等。每种多聚核糖体所包含的核糖体数量是由mRNA的长度来决定的,也就是说,mRNA越长,合成的多肽分子量越大,核糖体的数目也越多。Waner和Rich等发现网织红细胞内合成血红蛋白分子的多聚核糖体常常含有5个核糖体。根据血红蛋白的一条多肽链的大小(约150个氨基酸)可推算出其mRNA分子的长度约为150nm。他们用密度梯度离心技术与电镜负染色技术相结合,观察到网织红细胞内,多聚核糖体是由一条直径约1~1.5nm的mRNA串连5个(有时6个或4个)核糖体,相邻核糖体的中心间距为30~35nm,多聚核糖体的总长约150nm,这与前面的推论相符。细菌的β半乳糖苷酶由1 100个氨基酸残基组成,它的多聚核糖体中含有约50个核糖体,如将β半乳糖苷酶的基因截短,mRNA的长度随之缩短,多聚核糖体的大小及核糖体的数目也成比例地减少。在真核细胞中每个核糖体每秒能将两个氨基酸残基加到多肽链上,而在细菌细胞中可将20个氨基酸加到多肽链上,因此合成一条完整的多肽链平均需要20秒~几分钟,即使在这样短的时间里,当第一个核糖体结合到mRNA上起始蛋白质合成后,不久第二个核糖体便结合到mRNA上,相邻的核糖体间距约80个核苷酸的距离。由于蛋白质的合成是以多聚核糖体的形式进行,这样细胞内各种多肽的合成,不论其分子量的大小或是mRNA的长短如何,单位时间内所合成的多肽分子数目都大体相等,即在相同数量的mRNA的情况下,可大大提高多肽合成速度, 特别是对于分子量较大的多肽。多肽合成速度提高的倍数与结合在mRNA上的核糖体数目成正比。在细胞周期的不同阶段,细胞中数以万种的mRNA有些在合成,有些在降解,其种类与浓度不断发生变化,以多聚核糖体的形式进行多肽合成,对mRNA的利用及对其数量的调控更为经济和有效。原核细胞中,在mRNA合成的同时,核糖体就结合到mRNA上,即由DNA转录mRNA和由mRNA翻译成蛋白质是同时并几乎在同一部位进行,所分离的多聚核糖体常常与DNA结合在一起。真核细胞中,多聚核糖体或附着在内质网上,或游离在细胞质基质中。一些证据表明游离的多聚核糖体结合在细胞骨架上。
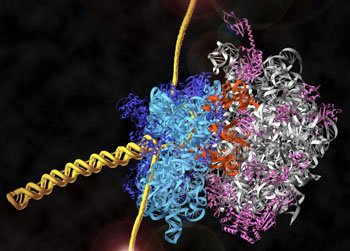
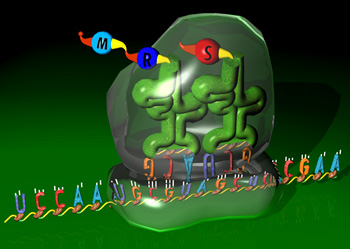
细胞中最主要的三种RNA―mRNA、rRNA和tRNA通过核糖体结合在一起,共同完成遗传信息表达中的最后一步――蛋白质的翻译。证明rRNA具有肽酰转移酶的活性,大大提高对探索生命起源中最基本也是最富有挑战性的问题――细胞遗传信息装置起源的兴趣。生命是自我复制的体系。推测最早出现的简单生命体中的生物大分子,应是既具有遗传信息载体功能又具有酶的催化功能。当今细胞遗传信息传递装置中的DNA,RNA和蛋白质三种生物大分子,DNA仅具有信息载体功能,而无酶活性;蛋白质具有多种酶活性而尚未发现有遗传信息载体功能;只有RNA既具有信息载体功能又具有酶的催化功能。因此,推测RNA可能是生命起源中最早的生物大分子。RNA的信息功能人们早已明了,mRNA即为信使RNA。许多病毒如爱滋病病毒和流感病毒等基因载体就是RNA,故称RNA病毒。但RNA催化功能是近二十年才被发现。随后人们发现一系列具有催化作用的RNA统称核酶(ribozyme)。核酶不仅可催化RNA和DNA水解、连接、mRNA的拼接(splicing),在体外已证明某些RNA还可催化RNA聚合反应以及RNA的磷酸化、氨酰基化和烷基化等多种反应。核糖体rRNA中可能具有肽酰转移酶的活性,在蛋白质合成中起着关键作用。从化学性质上推测,核糖核酸的基本成份―核糖,很容易由当时地球表面含量丰富的甲烷来合成。而脱氧核糖则需要经核糖还原而成,这一反应在细胞中是由专一的酶完成的。因此,生命的最早形式可能是由膜围绕的一套具有自我复制能力的分子体系和简单的物质与能量供应体系,其遗传物质的载体是RNA而不是DNA。RNA的催化效率远远低于蛋白质,因此整个体系复制效率很低。在漫长的进化过程中,由RNA催化产生了蛋白质,进而DNA代替了RNA的遗传信息功能,蛋白质则取代了绝大部分RNA酶的功能,逐渐演化成今天的细胞。由于DNA链比RNA链稳定,双链比单链稳定且DNA链中胸腺嘧啶代替了RNA链中的尿嘧啶,使之易于修复,作为遗传物质载体则有可能贮存大量的信息并能更稳定地遗传。由于蛋白质结构的多样性与构象的多变性,不仅比RNA更为有效地催化多种反应,而且也提供更为复杂的细胞结构成分。这也是当今结构和功能复杂的细胞进化的基础。很有趣的是至今在遗传信息表达体系中,不仅还要通过RNA完成遗传信息传递和密码的翻译,而且一些重要的反应过程如mRNA的拼接和蛋白质的合成仍需RNA的催化作用。