细胞膜细胞核 线粒体叶绿体内质网 核糖体高尔基体 溶酶体微体微丝微管 中间纤维
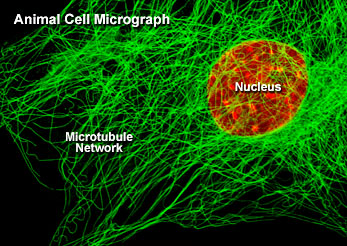
微管
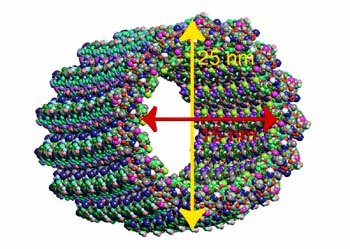
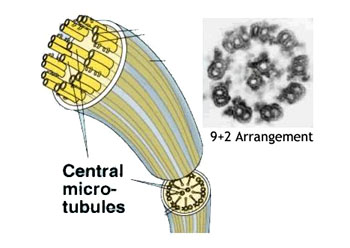
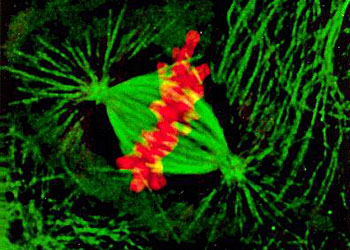
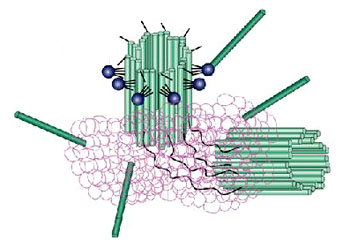
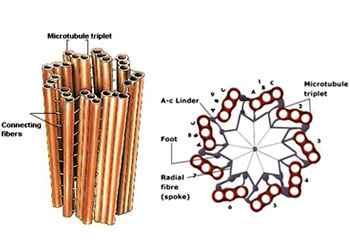
所有微管遵循同一原则由相似的蛋白亚基装配而成,主要装配方式是:首先,α-微管蛋白和β-微管蛋白形成长度为8nm的αβ二聚体,αβ二聚体先形成环状核心(ring),经过侧面增加二聚体而扩展为螺旋带,αβ二聚体平行于长轴重复排列形成原纤维(protofilament)。当螺旋带加宽至13根原纤维时,即合拢形成一段微管。新的二聚体再不断加到这一端微管的端点使之延长。最终微管蛋白与微管达到平衡。原纤维中重复的亚单位是αβ异二聚体,αβ→αβ→αβ,微管中这种亚单位排列即构成微管的极性,所有的微管都有确定的极性。微管的两个末端在结构上不是等同的,这是非常重要的结构特征。细胞内所有由微管构成的亚细胞结构也是有极性的。αβ→αβ即为头→尾的方向,微管蛋白加上或释放主要发生于(+)极,微管的延长主要依靠在(+)极组装GTP-微管蛋白,然后GTP水解为GDP或GTP与微管蛋白分离。目前的微管装配动态模型认为,微管两端具GTP帽(取决于微管蛋白浓度),微管将继续组装,反之,具GDP帽则解聚。在一定条件下,微管一端发生装配使微管延长,而另一端发生去装配而使微管缩短,称为踏车现象。微管蛋白的合成是自我调节的,多余的微管蛋白单体结合于合成微管蛋白的核糖体上,导致微管蛋白mRNA降解。微管在体内的装配和去装配在时间和空间上是高度有序的,间期细胞中,细胞质微管与微管蛋白亚单位库处于相对平衡状态。有丝分裂期中,胞质微管装配和去装配动态受细胞周期调控,发生显著改变,分裂前期,胞质微管网络中的微管去装配,游离的微管蛋白亚单位组装为纺锤体。分裂末期,发生逆向转变。此外,细胞中存在一些非常稳定的微管结构,如纤毛,鞭毛等。微管在生理状态及实验处理解聚后重新装配的发生处称为微管组织中心(microtubule-organizing center,MTOC)。动物细胞的MTOC为中心体。MTOC决定了细胞微管的极性,微管的(-)极指向MTOC,(+)极背向MTOC。体外微管装配条件:(1)微管蛋白浓度:随温度和技术条件而异,有一定的临界浓度,低于此浓度则不发生微管装配,大约为1mg/mL;(2)最适pH:pH6.9;(3)离子:Ca2+应尽可能除去,Mg2+为装配所必需;(4)温度:37℃微管蛋白二聚体装配成微管,0℃微管解聚为二聚体。