细胞膜
细胞核
线粒体
叶绿体
内质网
核糖体
高尔基体
溶酶体
微体
微丝
微管
中间纤维
你的位置:
首页
>
细胞
> 减数分裂
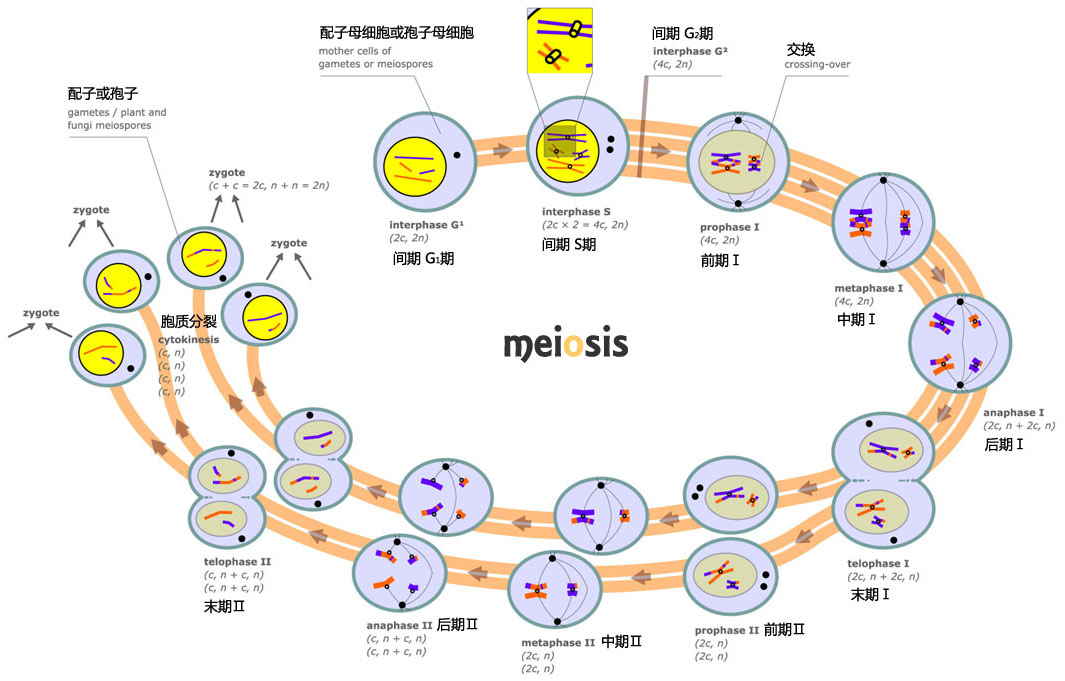
减数分裂(meiosis)是一种特殊的有丝分裂形式,仅发生于有性生殖细胞形成过程中的某个阶段。减数分裂的主要特点是,细胞仅进行一次DNA复制,随后进行两次分裂。两次分裂分别称为减数分裂期I和减数分裂期II。在两次分裂之间,还有一个短暂的分裂间期。两性生殖细胞经过减数分裂,各自的染色体数减少一半。再经过受精,形成合子,染色体数恢复到体细胞的染色体数目。减数分裂的意义在于,即有效地获得父母双方的遗传物质,保持后代的遗传性,又可以增加更多的变异机会,确保生物的多样性,增强生物适应环境变化的能力。相反,假如在有性生殖过程中没有减数分裂,生殖细胞染色体数不能减半,经过受精,其染色体数必将倍增。细胞体积也会相应增加,生物个体体积也会增长。代代相传,其生命活动将无法适应环境变化,终将受到自然淘汰。因而,减数分裂是生物有性生殖的基础,是生物遗传、生物进化和生物多样性的重要基础保证。与有丝分裂相似,在减数分裂之前的间期阶段,也可以人为地划分为G1期,S期,G2期等三个时期。但此间期阶段也有其鲜明的特殊性。为区别于一般的细胞间期,常把减数分裂前的细胞间期称为减数分裂前间期(premeiotic interphase)。
减数分裂
Meiosis
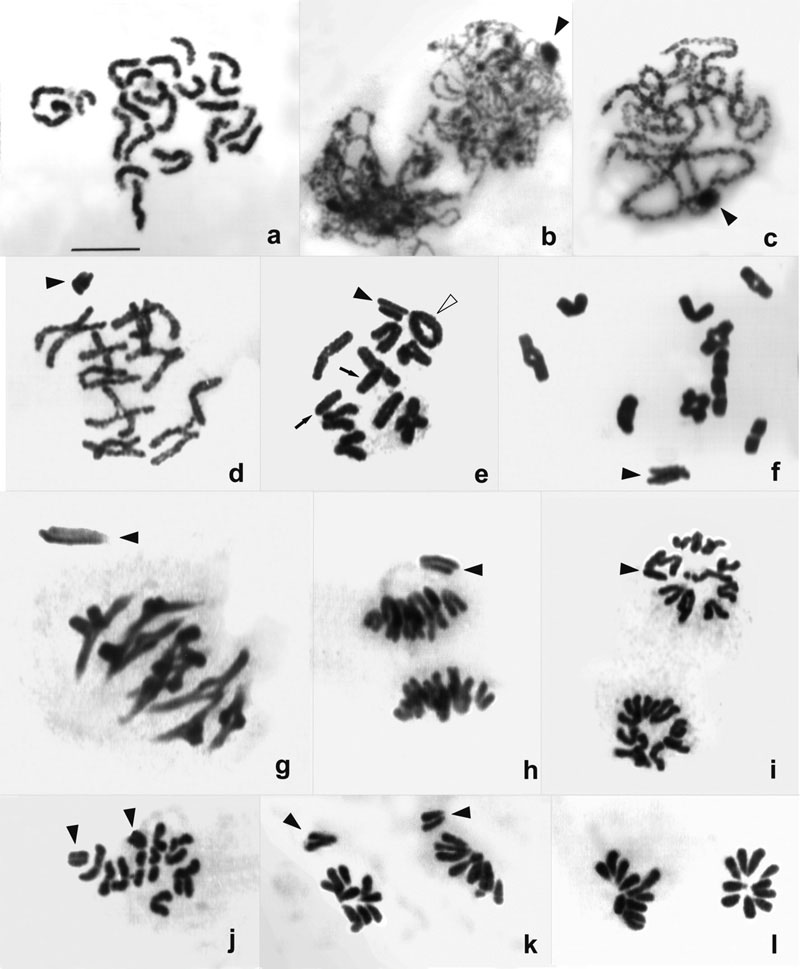
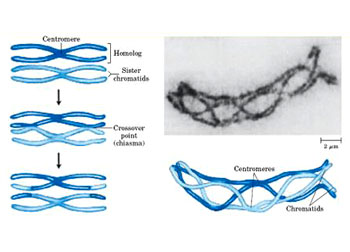
一种热带狼蛛的精原细胞减数分裂:a. 前间期;b. 偶线期;c. 粗线期;d. 早期双线期;e. 双线期结束;f. 终变期;g. 中期I;h. 后期I;i. 末期I;j. 中期II;k. 后期II;l. 末期II (Bar=10μm)
减数分裂前间期
减数分裂前间期(premeiotic interphase)的最大特点在于其S期持续时间较长,同时也发生一系列与减数分裂相关的特殊事件。例如,蝾螈(Triturus)体细胞有丝分裂前S期约为12小时,而减数分裂前S期则可持续10天。小鼠有丝分裂前S期约为5~6小时,而其减数分裂前S期约为14小时。另一个重要特点是,在植物百合中发现,其减数分裂前间期的S期仅复制其DNA总量的99.7~99.9%,而剩下的0.1~0.3%要等到减数分裂前期阶段才进行复制。科学家发现,这些推迟复制的DNA被分割为5 000~10 000个小片段,分布于整个基因组中,每个小片段长约1 000~5 000个碱基对。另外还发现,有一种蛋白质,称为L蛋白,在减数分裂前间期与上述DNA小片段结合,阻止其复制。这些DNA小片段被认为与减数分裂前期染色体配对和基因重组有关。大多数生物,其减数分裂前间期的细胞核大于其体细胞核。染色质也多凝集成异染色质。这种变化的意义虽不明了,但一般认为与染色体配对和基因重组有关。另外,根据生物种类不同,减数分裂前间期的G2期的长短变化较大。有的G2期短,有的则和有丝分裂前间期的G2期长短相当,也有的可以在G2期停滞较长一段时间,直到受到新的刺激来打破这种停滞。由减数分裂前G2期细胞进入两次有序的细胞分裂,即第一次减数分裂和第二次减数分裂。两次减数分裂之间的间期或长或短,但无DNA合成。
减数分裂期I
减数分裂期I(meiosisI)与体细胞有丝分裂期,有许多相似之处。其过程也可以人为地划分为前期I,前中期I,中期I,后期I,末期I和胞质分裂I等六个阶段。但减数分裂期I又有其鲜明的特点。其主要表现在分裂前期的染色体配对和基因重组以及其后的染色体分离方式等方面。
前期I
前期I(prophaseI)持续时间较长。在高等生物,其时间可持续数周,数月,数年,甚至数十年。在低等生物,其时间虽相对较短,但也比有丝分裂前期持续的时间长得多。在这漫长的时间过程中,要进行染色体配对和基因重组。此外,也要合成一定量的RNA和蛋白质。根据细胞形态变化,又可以将前期I人为地划分为细线期,偶线期,粗线期,双线期,终变期等五个阶段。
细线期又称凝集期,染色体发生凝集。染色体已经进行了复制,一条染色体应由两条染色单体组成,但一般看不出两条染色单体,好像单股。细线的两端通过接触斑与核膜相连,核体积逐渐增大。
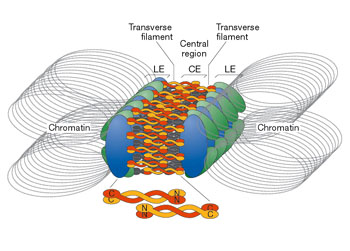
偶线期又称配对期,同源染色体两两配对,即联会。在配对部位,沿两条同源染色体间长轴形成一特殊结构――联会复合体,包括侧生组分、中间区、中央组分三部分。
粗线期又称重组期,联会的两条同源染色体紧密结合,同源的非姐妹单色单体间发生局部交换,形成新的等位基因组合。此时每对配对的染色体中含有四条染色单体,称为四分体。
细线期
(leptotene,leptonema):为前期I的开始阶段。首先发生染色质凝集,染色质纤维逐渐折叠,螺旋化,变短变粗,包装成在显微镜下可以看到的细纤维样染色体结构。因而,有人将细线期也称为凝集期(condensation stage)。细线期与有丝分裂前期起始阶段也有着明显的不同。首先,在细线期染色质虽然发生凝集,但两条染色单体的臂并不分离,在显微镜下看不到双线样染色体结构,而是呈细的单线状。有人认为,将两条染色单体紧密联系在一起的一种因素可能是在减数分裂前S期未被复制的DNA片段。由于DNA复制尚未完成,两条染色单体不能完全形成,因而,染色单体的臂也就不能分离。另一个明显不同点是,在细纤维样染色体上,出现一系列大小不同的颗粒状结构,称为染色粒(chromomere)。虽然已经知道染色粒由染色质组成,但其功能并不清楚。细线期还有一个明显的特点,即染色体端粒(telomere)通过接触斑与核膜相连。而染色体的其它部分以袢状伸延到核质中。有的物种,接触斑位于细胞核的一侧,染色体呈花束状向核内其它部位伸延。因而,有人也将细线期称为花束期。
偶线期
(zygotene,zygonema):主要发生同源染色体配对(pairing),即来自父母双方的同源染色体逐渐靠近,沿其长轴相互紧密结合在一起。因而,偶线期又称为配对期(pairing stage)。配对过程是专一性的,仅发生于同源染色体之间,非同源染色体之间不进行配对。关于同源染色体之间相互识别的机制,目前尚不清楚。配对以后,两条同源染色体紧密结合在一起所形成的复合结构,称为二价体(bivalent)。由于每个二价体由两条染色体构成,共含有四条染色单体,因而又称为四分体(tetrad)。但此时的四分体结构并不清晰可见。同源染色体配对的过程称为联会(synapsis)。联会初期,同源染色体端粒与核膜相连的接触斑相互靠近并结合。从端粒处开始,这种结合不断向其它部位伸延,直到整对同源染色体的侧面紧密联会。联会也可以同时发生在同源染色体的几个点上。在联会的部位形成一种特殊复合结构,称为联会复合体(synaptonemal complex)。联会复合体沿同源染色体长轴分布,宽约1.5~2μm,在电镜下可以清楚地显示其细微结构。联会复合体被认为与同源染色体联会和基因重组有关。在偶线期发生的另一个重要事件是合成在S期未合成的约0.3%的DNA(偶线期DNA,即zygDNA)。若用DNA合成抑制剂抑制zygDNA合成,联会复合体的形成将受到抑制。zygDNA在偶线期转录活跃。转录的RNA被称为zygRNA。zygDNA转录也被认为与同源染色体配对有关。
联会(synapsis)也称配对,指细胞减数分裂前期I的偶线期,来自父本和母本的同源染色体,两两靠拢进行准确配对,形成四分体,即一对染色体含有四条染色单体,但仅有两个着丝点。联会是减数分裂的重要特征。
联会复合体(synaptonemal complex, SC)是减数分裂偶线期两条同源染色体之间形成的一种结构,它与同源染色体的配对、交换和分离密切相关。联会复合体沿同源染色体长轴分布,宽约1.5~2μm,在电镜下可以清楚地显示其细微结构。
联会复合体(synaptonemal complex, SC)是减数分裂偶线期两条同源染色体之间形成的一种结构,它与同源染色体的配对、交换和分离密切相关。联会复合体沿同源染色体长轴分布,宽约1.5~2μm,在电镜下可以清楚地显示其细微结构。
粗线期
(pachytene,pachynema):开始于同源染色体配对完成之后。这一过程可以持续几天至几个星期。在此过程中,染色体进一步浓缩,变粗变短,并与核膜继续保持接触。同源染色体仍紧密结合,并发生等位基因之间部分DNA片段的交换和重组,产生新的等位基因的组合。此时在联会复合体部位的中间,出现一个新的结构,呈圆球形,椭球形或长约0.2mm的棒状,称为重组节(combination nodule)。重组节直径约90nm,内含蛋白质等成分,结构尚不清楚。重组节可能直接参与基因重组。有些生物,在整个减数分裂过程中不出现重组节,因此中并无基因重组发生。在粗线期,也合成一小部分尚未合成的DNA,称为P-DNA。P-DNA大小约为100~1 000bp,编码一些与DNA点切(nicking)和修复(repairing)有关的酶类。粗线期另一个重要的生化活动是,合成减数分裂期专有的组蛋白,并将体细胞类型的组蛋白部分或全部地置换下来。这种置换也许在一定程度上参与了基因重组过程,或反映出减数分裂前期染色体结构的变化。在许多动物的卵母细胞发育过程中,粗线期还要发生rDNA扩增。即编码rRNA的DNA片段从染色体上释放出来,形成环形的染色体外DNA,游离于核质中,并进行大量复制,形成数千个拷贝的rDNA。如在非洲爪蟾(Xenopus)卵母细胞中,经过rDNA扩增,可以产生大约2500个拷贝的rDNA。这些rDNA将参与形成附加的核仁,进行RNA转录。
双线期
(diplotene,diplonema):重组阶段结束,同源染色体相互分离,仅留几处相互联系。同源染色体的四分体结构变得清晰可见。同源染色体仍然相联系的部位称为交叉(crossover,chiasma)。交叉的数量变化不定。即使在同种物种的不同细胞之间,交叉的数量也不相同。一般而言,在每个染色体臂上至少有一个交叉。在电镜下可见,交叉部位含有残留的联会复合体结构。许多动物在双线期阶段,同源染色体或多或少地要发生去凝集,RNA转录活跃。关于染色体去凝集的程度,有的种类低到不易觉察,有的种类则高到几乎与一般间期细胞相似。在许多动物,尤其是鱼类,两栖类,爬行类和鸟类的雌性动物,染色体去凝集成一种特殊的巨大染色体结构,形状好象油灯的灯刷,称为灯刷染色体(lampbrush chromosome)。在灯刷染色体上有许多侧环结构,是进行活跃转录RNA的部位。RNA转录,蛋白质翻译以及其它物质的合成等,是双线期卵母细胞体积增长所必需的(见第八章)。目前比较清楚,在灯刷染色体侧环上合成的RNA主要为前体mRNA。前体mRNA合成以后,很快被剪辑为mRNA。有些种类的mRNA,如编码组蛋白、核糖体蛋白和卵黄蛋白的mRNA很快会在细胞质中翻译为蛋白质。许多其它种类的mRNA则和蛋白质结合,以非活跃形式储备在卵母细胞质中。直到卵细胞成熟并受精以后,这些储备的mRNA才能转变为活跃状态,进行蛋白质翻译。在灯刷染色体一定的侧环上,也可以检测到tRNA和5SrRNA的转录。双线期持续时间一般较长,其长短变化很大。两栖类卵母细胞的双线期可持续将近一年,而人类的卵母细胞双线期从胚胎期的第五个月开始,短者可持续十几年,到性成熟期开始;长者可达四、五十年,到生育期结束。
终变期
(diakinesis):染色体重新开始凝集,形成短棒状结构。如果有灯刷染色体存在,其侧环回缩,RNA转录停止,核仁消失,四分体较均匀地分布在细胞核中。同时,交叉向染色体臂的端部移行。此移行过程称为端化(terminalization)。到达终变期末,同源染色体之间仅在其端部和着丝粒处相互联结。终变期的结束标志着前期I的完成。
双线期又称合成期。联会的同源染色体分离,但尚未完全分开,仍有若干处相连。这些相连点式染色体间发生互换的结果。染色体与核被膜脱离接触。mRNA和rRNA转录十分旺盛。
终变期又称再凝集期,染色体凝集成短棒状,核仁开始消失,核膜开始解体。四分体较均匀地分布在核中,同源染色体间依靠端部交叉相结合,姐妹染色体由着丝粒相连。
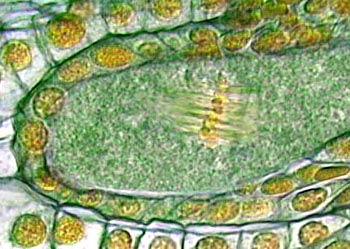
中期I时,核膜破裂,纺锤体形成,配对的同源染色体(二价体)在纺锤丝的作用下,移到细胞的赤道面上。与有丝分裂不同的是,每个四分体含有四个动粒(图为孢子减数分裂的中期I)
中期I
前期结束,细胞逐渐转入分裂中期I(metaphaseI)。在此过程中,要进行纺锤体组装。纺锤体形成过程和结构与一般有丝分裂过程中的相类似。到核膜破裂,标志着中期I的开始。纺锤体微管侵入核区,捕获分散于核中的四分体。四分体逐渐向赤道方向移动,最终排列在赤道面上。与有丝分裂不同的是,每个四分体含有四个动粒。其中一条同源染色体的两个动粒位于一侧,另一条同源染色体的两个动粒位于另一侧。从纺锤体一极发出的微管只与一个同源染色体的两个动粒相连,从另一极发出的微管也只与另一个同源染色体的两个动粒相连。
后期I
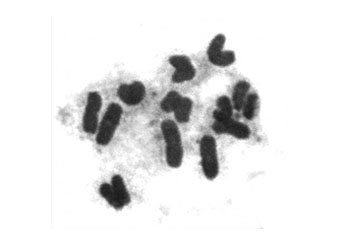
同源染色体对相互分裂并向两极移动,标志着后期I(anaphaseI)的开始。移向两极的同源染色体均是含有两条染色单体的二倍体。其结果是,到达每一极的染色体的数量为细胞内染色体总数量的一半。另外,各四分体之间同源染色体向两极移动是一个随机的过程,因而到达两极的染色体会出现许许多多的排列方式。如人类细胞有23对染色体,从理论上讲将会产生223种不同的排列方式。如此庞大的排列方式,即使不发生基因重组,得到遗传上完全相同的配子机率也只有八百四十万分之一。再加上基因重组和精子与卵子的随机结合,要想得到遗传上完全相同的个体几乎是不可能的,除非是同卵双生个体,其遗传性状可能相同。
末期I、胞质分裂I和减数分裂间期
经过后期I后,细胞进一步的变化主要有两种类型:第一种类型,染色体到达两极,并逐渐进行去凝集。在染色体的周围,核被膜重新组装,形成两个子细胞核。同时,随着染色体分离并向两极移动,细胞质也开始分裂,完全形成两个间期子细胞。此时的间期细胞虽具有一般间期细胞的基本结构特征,但又有着重要区别,即它们不再进行DNA复制,也没有G1、S和G2期之分。间期持续时间一般较短,有的仅作短暂停留。为区别于一般细胞间期,特将其称为减数分裂间期(interkinesis);第二种类型,即细胞进入末期后,不是完全回复到间期阶段,而是立即准备进行第二次减数分裂。
第二次减数分裂
第二次减数分裂过程与有丝分裂过程非常相似。即经过分裂前期II、中期II、后期II、末期II和胞质分裂II等几个过程。每个过程中细胞形态变化也与有丝分裂过程相似。对于上述第二种类型,染色体到达两极后,减数分裂I的纺锤体去组装,两极的中心粒和星体,此时一分为二,重新组装成两个纺锤体。染色体在原来两极的位置重新排列,形成新的赤道板。此时即为中期II。此后的发展则与一般有丝分裂相似。经过第二次减数分裂,共形成四个子细胞。但它们以后的命运随生物种类不同而不同。在雄性动物,四个细胞大小相似,称为精子细胞,将进一步发展成为四个精子。在雌性动物,第一次分裂为不等分裂,即第一次分裂后产生一个大的卵母细胞和一个小的极体,称为第一极体。第一极体将很快死亡解体,有时也会进一步分裂为两个小细胞,但没有功能。卵母细胞将继续进行第二次减数分裂,也为不等分裂。其结果是产生一个卵细胞和一个第二极体。第二极体也没有功能,很快解体。因此,雌性动物减数分裂仅形成一个有功能的卵细胞。高等植物减数分裂与动物减数分裂类似,即雄性产生四个有功能活性的精子,而雌性仅产生一个有功能活性的卵细胞。
减数分裂(meiosis)是性细胞连续进行两次核分裂,而染色体只复制一次,由此产生四个单倍体细胞(配子),染色体数目减半(2n→n)的特殊细胞分裂方式
性染色体的分离
不同性别之间,其性染色体构成不同。绝大多数雌性动物细胞的两条性染色体构成为XX,而雄性细胞性染色体为XY或XO(即无Y染色体)。也有一些种类正好相反,雄性为XX两条染色体,雌性为XY两条染色体。对于含有两条XX性染色体的细胞,两条XX染色体象常染色体一样进行正常配对、交换和分离。而含有XY性染色体的细胞,两条性染色体的形态结构不同,基因含量也不同。在前期I,二者是如何配对和分离的呢?一般讲,有些物种的XY染色体间可能会含有一些同源区段,有的可能不含任何同源区段。对于含有同源区段的XY染色体,如人类的XY染色体,在前期I可以进行配对。不管XY染色体配对与否,二者都将和常染色体一样,在分裂中期I排列到赤道面上。其后,随常染色体分离而相互分离,并各自移向两极。到第二次减数分裂,XY染色体和常染色体一样,其两条染色单体再进行分离。偶尔也可出现XY染色体的染色单体在第一次减数分裂时就相互分离的现象,致使产生的两个细胞各含有一个X染色单体和一个Y染色单体。到第二次减数分裂时,每个细胞的X和Y染色单体再分配到两个细胞中。对于XO物种(主要是昆虫),在第一次减数分裂时,X染色体移向一极。结果将产生一个含X染色体的细胞和一个不含性染色体的细胞。到第二次减数分裂,含X染色体的细胞分裂为两个含X染色单体的细胞;不含性染色体的细胞也一分为二,形成两个不含性染色体的细胞。偶尔也可以看到X染色体的两个染色单体在第一次减数分裂时即相互分离,产生两个各含一个X染色单体的细胞。到第二次减数分裂时,X染色单体仅分配到一个细胞中。其最终结果是,一个XO细胞经过减数分裂,产生两个含X染色体的细胞和两个无性染色体的细胞。